
North American coral recovery after the end-Triassic mass extinction, New York Canyon, Nevada, USA
Montana S. Hodges and George D. Stanley Jr.
In this article
Full Text View
Volume 25 Issue 10 (October 2015)
GSA Today
Article, pp. 4-7 | Abstract | PDF (530KB)
North American coral recovery after the end-Triassic mass extinction, New York Canyon, Nevada, USA
[Table: Table of Contents Introduction Geologic Setting and Biochronology of New York Canyon Jurassic R...]
Montana S. Hodges* and George D. Stanley Jr.
University of Montana Paleontology Center, 32 Campus Drive, Missoula, Montana 59812, USA
ABSTRACT
A Triassic-Jurassic (T/J) mass extinction boundary is well represented stratigraphically in west-central Nevada, USA, near New York Canyon, where the Gabbs and Sunrise Formations contain a continuous depositional section from the Luning Embayment. The well-exposed marine sediments at the T/J section have been extensively studied and reveal a sedimentological and paleontological record of intense environmental change and biotic turnover, which has been compared globally. Unlike the former Tethys region, Early Jurassic scleractinian corals surviving the end-Triassic mass extinction are not well-represented in the Americas. Here we illustrate corals of Early Sinemurian age from Nevada located at three horizons above the T/J boundary. These well-preserved corals represent one of the earliest Jurassic appearances in North America and the earliest in the United States. Their co-occurrence with bivalves, gastropods, and ammonites adds additional faunal elements to the study. The corals are exclusively solitary and occur in profusion packed within beds. They all belong to the family Stylophyllidae, known to have been extinction resistant. These post-extinction corals support the Hispanic Corridor hypothesis and provide new data on biotic recovery following the end-Triassic mass extinction.
Email:
Manuscript received 6 Mar. 2015; accepted 9 July 2015.
doi: 10.1130/GSATG249A.1
Introduction
Mass extinction events punctuate the evolution of marine environments, and recovery biotas paved the way for major biotic changes. Understanding the responses of marine organisms in the post-extinction recovery phase is paramount to gaining insight into the dynamics of these changes, many of which brought sweeping biotic reorganizations. One of the five biggest mass extinctions was that of the end-Triassic, which was quickly followed by phases of recovery in the Early Jurassic. The earliest Jurassic witnessed the loss of conodonts, severe reductions in ammonoids, and reductions in brachiopods, bivalves, gastropods, and foraminifers. Reef ecosystems nearly collapsed with a reduction in deposition of CaCO3. Extensive volcanism in the Central Atlantic Magmatic Province and release of gas hydrates and other greenhouse gases escalated CO2 and led to ocean acidification of the end-Triassic (Hautmann et al., 2008).
Reef-building scleractinian corals and spinctozoid sponges experienced severe setbacks after the Late Triassic reef optimum (Stanley, 2003; Flügel, 2002). Although coral recovery began soon after the extinction (Lathuilière and Marchal, 2009), their diversity was low (Fig. 1). Compared to other calcified biotas, reef-building corals and sponges experienced proportionately greater losses (corals, 96.1%; sponges 91.4%), possibly related to their reduced physiological control of calcification with respect to aragonite saturation (Hautmann et al., 2008). Most reports of Early Jurassic corals come from tropical to subtropical regions of the former Tethys, now comprising central Eurasia and Morocco (Lathuilière and Marchal, 2009). Jurassic examples from the Americas are rare, while Triassic corals are relatively common (Stanley, 1997).
Figure 1
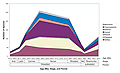
Reef diversity through time plotted by highest to lowest corallite integration levels. Information derived from Paleobiology Database; figure from Shepherd, 2013. Het—Hettangian.
Because Early Jurassic corals from North America are so rare, new occurrences are vital to understanding biotic responses in the post–mass extinction interval. Here we make a preliminary report of the earliest Jurassic corals from the USA and some of the earliest in North America. These examples occur at New York Canyon in west-central Nevada’s Sunrise Formation (Fig. 2). The site has attracted international attention because it is among the best documented T/J sections (Lucas et al., 2007). The coral occurrences in this section are near the T/J boundary, making the Nevada site ideal for understanding the dynamics of coral recovery and comparing them with the Tethys.
Figure 2
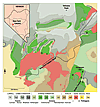
Geologic map of New York Canyon, Nevada, USA. The Involutum Zone coral-collecting sites labeled A–I are located in the upper layers of the Js2 unit. Map modified from Ferguson and Muller, 1949.
Geologic Setting and Biochronology of New York Canyon
The Gabbs and Sunrise Formations at New York Canyon were first described by Muller and Ferguson (1936) and have become an international reference section for the T/J boundary. In New York Canyon, the Lower Jurassic Sunrise Formation and underlying Upper Triassic Gabbs Formation represent coherent and conformable alternating siltstone and limestone marine sequences deposited in the shallow portion of the Luning Embayment. Investigations on the stratigraphy (Taylor et al., 1983; Hallam and Wignall, 2000), the biochronology (Guex, 1995; Taylor et al., 2001), and carbon isotopes (Guex et al., 2004; Ward et al., 2007; Porter et al., 2014) have established the Ferguson Hill section as a relatively complete marine sequence spanning the Upper Triassic to Lower Jurassic boundary (Guex et al., 1997; McRoberts et al., 2007; Lucas et al., 2007; Ritterbush et al., 2014).
Early Jurassic corals forming the basis of this report were collected from nine sites in the Ferguson Hill Member of the Sunrise Formation (in the vicinity of New York Canyon, Fig. 2) including the Involutum Zone Type Section (Taylor et al., 2001) and the Ferguson Hill section, just above the T/J boundary (Fig. 3). All locations are in the Early Sinemurian Involutum Zone, a Cordilleran ammonite zone correlated with the upper Bucklandi subzone to lower Semicostatum zone of northwestern Europe (Taylor et al., 2001).
Figure 3

East side of Ferguson Hill, New York Canyon, Nevada. Coral-bearing Involutum Zone is visible at the top of the photo where resistant darker siliceous limestone beds are overlain by primarily non-resistant light brown siltstone beds 40 m above the T/J boundary.
The Involutum Zone at New York Canyon corresponds to the 15-m-thick, thin-to-medium bedded limestone and siltstone superjacent to the more thickly bedded dark gray-to-black chert-rich limestone (Taylor et al., 1983). The base of the New York Canyon Involutum Zone is well-demarcated by bioclastic limestone overlaying more resistant, darker siliciclastic limestone beds (Taylor et al., 2001) and is very pronounced at Ferguson Hill. The Involutum Zone occurs in the upper beds of the Js2 unit (Fig. 2).
Corals in the New York Canyon area are biostratigraphically constrained by ammonites in three horizons measured above the base of the Involutum Zone. The first corals are at 4 m in the Coroniceras fergusoni horizon. The next corals occur at 10–12 m in the Volcanoense Subzone, and lastly at 13–15 m in the Mullerense Subzone (Taylor et al., 2001). The most abundant coral-bearing outcrops are on top of Ferguson Hill at N 38°29.237'; W 118°5.033' and the first ridge southeast of the Involutum Zone Type Section at N 38°29.478'; W 118°5.669'. Co-occurring with the corals is Weyla alata, an endemic Hettangian to Pliensbachian bivalve of eastern Panthalassa (Damborenea and González-León, 1997). Corals at New York Canyon were first recorded by Muller and Ferguson (1936) and subsequently recognized by current researchers (Guex, personal commun., Sept. 2014; Caruthers, personal commun. Aug. 2014; Ritterbush et al., 2014). Unfortunately, no taxonomic determinations have been published pertaining to the corals.
Hundreds of solitary corals were collected. Genera identified are Stylophyllopsis, Protostylophyllum, and Haimeicyclus (Fig. 4); all belong to the family Stylophyllidae. They are enclosed in fine-grained, poorly sorted siliciclastic limestone with abundant coated and micritized molluscan bioclasts. Most coral calices are infilled with peloids, indicating a shallow, well-lit, energetic marine environment. While a few corals occur in life positions, many appear overturned and reworked (Fig. 5). The early Sinemurian stylophyllids at New York Canyon are unique in being solitary, with gregarious associations. The coral larvae were likely fixosessile, attaching either to small grains or shell fragments. They also show distinctive expansions and constrictions, expressed both internally and externally on the epithecal wall (Fig. 4). This indicates polyp rejuvenation, likely as an ecological avoidance to smothering by frequent, perhaps cyclic influxes of exogenous sediment. Such cyclic rejuvenescence is well known among solitary rugose (Berkowski, 2012) and modern corals (Chevalier and Beauvais, 1987). Corals at New York Canyon were capable of surviving smothering by both sediment rejection and polyp regeneration. The lack of colonial corals is unusual and might indicate that these solitary taxa were better adapted at sediment rejection or that they were ahermatypic and, like many living examples, utilized a simple growth form.
Figure 4

Cross-sectional views of coral from the Ferguson Hill Member of the Sunrise Formation. Haimeicyclus (left; specimen no. UMPC14687), Protostylophyllum (middle; specimen no. UMPC14688). A weathered specimen of Stylophyllopsis (right; specimen no. UMPC14689) displays a series of expansions and constrictions of the corallite indicating high sedimentation rates.
Figure 5

In-situ corals from the Ferguson Hill Member displaying reworked and life positions. Scale bar: 1 cm. Specimen no. UMPC14690.
Jurassic Recovery Among Corals
Many marine faunas underwent an Early Jurassic post-extinction recovery including corals. Earliest Jurassic (Hettangian) corals occur in mostly low diversity associations in the former Tethys region and appear absent in North America. Sinemurian coral deposits are known in reefs of British Columbia, Canada (Stanley and Beauvais, 1994), and Sonora, Mexico (González-León, et al., 2005). Sinemurian patch reefs also come from Peru (Wells, 1953). Non–reef building Hettangian to Sinemurian corals also occur in Peru (Senowbari-Daryan and Stanley, 1994) and Chile (Prinz, 1991). The Early Jurassic corals of the Americas are mostly holdover taxa from the Triassic.
Tethyan reef recovery started in the Hettangian (Kiessling et al., 2009; Gretz et al., 2013) but coral diversity was low. From the standpoint of changes in taxonomic composition, Tethyan corals show a recovery within the post-extinction survival interval (Beauvais, 1989). Their taxonomic composition reveals dominantly Triassic holdover taxa during the earliest Hettangian to Sinemurian intervals with increasingly new taxa into the Pliensbachian. By the Toarcian, an entirely new Jurassic coral assemblage evolved with few Triassic holdovers (Beauvais, 1986). From the standpoint of coral and reef diversity, it was not until the Middle Jurassic that the ecosystem was fully recovered (Stanley, 1997). By that time most Triassic holdovers were extinct.
Corals of the family Stylophyllidae (including New York Canyon) survived the end-Triassic extinction in the Tethys and diversified during the Early Jurassic (Melnikova and Roniewicz, 2012). This group is represented by solitary and phaceloid corals, which flourished briefly in the post-extinction aftermath. The branching Phacelostylophyllum rugosum, for example, was the reef builder in western Canada and Chile, while another Phacelostylophyllum species dominated the Early Jurassic reef in Peru (Stanley and Beauvais, 1994). In the Tethys, identical or closely related stylophyllids likewise constructed the two known earliest Jurassic patch reefs (Kiessling et al., 2009; Gretz et al., 2013). Stylophyllopsis, Protostylophyllum, and Haimeicyclus at New York Canyon reveal a strong taxonomic connection with Tethyan taxa. New York Canyon corals such as Protostylophyllum also reveal close affinities with Upper Triassic Tethyan species of the same genus (Roniewicz and Michalík, 2002).
Compared to the Tethys, precious little is known about American corals after the end-Triassic event. In west-central Nevada, Upper Triassic corals are well documented from the Luning and Osobb Formations (Roniewicz and Stanley, 2013). The Triassic corals of Nevada reveal strong North American endemism but strangely lack any representatives of the stylophyllids that are so pervasive in the Triassic Tethys. The cosmopolitan pattern of the Early Jurassic New York Canyon stylophyllid corals compared with those of the Tethys, along with the endemic pattern of the Late Triassic corals of Nevada, provide paleobiogeographic support for the Hispanic Corridor (Smith, 1983). The Hispanic Corridor was a narrow, embryonic Atlantic seaway hypothesized to have opened in Pliensbachian time, creating a shortcut connection between the Tethys and eastern Panthalassa (Aberhan, 2001). The New York Canyon corals indicate the existence of this seaway earlier in the Sinemurian, supporting the postulates of Sha (2002).
Discussion and Conclusions
The collapse of marine ecosystems at the end-Triassic is linked to ocean acidification, and a biocalcification crisis accounts for the scarcity of corals and reefs in the Early Jurassic (Martindale et al., 2012). The geographic proliferation of Late Triassic reefs through the Tethys is in stark contrast to their Early Jurassic reduction (Fig. 6). The Early Jurassic reef eclipse is revealed by only two rare examples, those in southern France (Kiessling et al., 2009) and Scotland (Gretz et al., 2013). This is the effect predicted for modern reefs under ocean acidification models projected 35–60 years from today (Hoegh-Guldberg et al., 2007). In Early Jurassic deposits of the Tethys, stylophyllid corals were common (Beauvais, 1976), along with cerioid Septastrea and Astrocoenia. Melnikova and Roniewicz (2012) reported Early Jurassic corals from the Pamir Mountains (Hettangian-Sinemurian) and noted how stylophyllids pass through the Triassic briefly proliferating during Early Jurassic time. A late Sinemurian coral reef reported from an outboard oceanic terrane in western Canada (Stanley and McRoberts, 1993) was constructed by large colonies of the stylophyllid Phacelostylophyllum (Stanley and Beauvais, 1994). A comprehensive database of Triassic and Jurassic Tethyan corals (Lathuilière and Marchal, 2009; Roniewicz and Morycowa, 1993) shows the survival pattern.
Figure 6
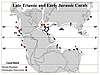
Paleogeographical map of Pangea (gray) showing reefs of the Late Triassic (blue boxes) and the Early Jurassic (red circles). The star represents Ferguson Hill Member corals. Data derived from the Palaeoreef database. Map modified from Lathuilière and Marchal, 2009.
The New York Canyon corals offer additional information on recovery in eastern Panthalassa along the craton of North America. Paleogeographically, the New York Canyon corals show a strong connection with Tethys but in contrast are exclusively solitary and exclusively stylophyllid taxa. Hettangian corals are unknown from North America, so for the present, the New York Canyon site may be the earliest North American Jurassic example. Analysis of these corals fills a neglected but important part of the T/J recovery phase in North America, while lending support for an earlier opening of the Hispanic Corridor.
REFERENCES CITED
- Aberhan, M., 2001, Bivalve palaeobiogeography and the Hispanic Corridor: time of opening and effectiveness of a proto-Atlantic seaway: Palaeo, v. 165, p. 375–394, doi: 10.1016/S0031-0182(00)00172-3.
- Beauvais, L., 1976, Révision des types de Madréporaires Liasiques décrits par Duncan (1867): Mémoires Société Géologique de France, v. 55, p. 1–84.
- Beauvais, L., 1986, Monographie des madréporaires du Jurassique inferieur du Maroc: Palaeontographica, v. 194A, p. 1–68.
- Beauvais, L., 1989, Jurassic corals from the circum Pacific area: Memoir of the Association of Australasian Palaeontologists, v. 8, p. 291–302.
- Berkowski, B., 2012, Life Strategies and Functions of Dissepiments in Rugose Coral Catactotoechus instabilis from the Lower Devonian of Morocco: Acta Palaeontologica Polonica, v. 57, p. 391–400, doi: 10.4202/app.2010.0048.
- Chevalier, J.P., and Beauvais, L., 1987, Ordre des scléractiniaires: Traité de zoologie: Anatomie, systématique, biologie, v. 3, p. 403–764.
- Damborenea, S.E., and González-León, C.M., 1997, Late Triassic and Early Jurassic bivalves from Sonora, Mexico: Revista Mexicana de Ciencias Geológicas, v. 14, p. 178–201.
- Ferguson, H.G., and Muller, S.W., 1949, Structural geology of the Hawthorne and Tonopah quadrangles, Nevada: U.S. Government Printing Office, U.S. Geological Survey Professional Paper 216, 55 p.
- Flügel, E., 2002, Triassic reef patterns, in Kiessling, W., Flügel, E., and Golonka, J., eds., Phanerozoic Reef Patterns: SEPM, v. 72, p. 391–463.
- González-León, C.M., Stanley, G.D., Jr., Gehrels, G.E., and Centeno-García, E., 2005, New data on the lithostratigraphy, detrital zircon and Nd isotope provenance, and paleogeographic setting of the El Antimonio Group, Sonora, Mexico, in Anderson, T.H., Nourse, J.A., McKee, J.W., and Steiner, M.B., eds., The Mojave-Sonora Megashear Hypothesis: Development, Assessment, and Alternatives: GSA Special Paper 393, p. 259–282.
- Gretz, M., Lathuilière, B., Martini, R., and Bartolini, A., 2013, The Hettangian corals of the Isle of Skye (Scotland): An opportunity to better understand the palaeoenvironmental conditions during the aftermath of the Triassic–Jurassic boundary crisis: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 376, p. 132–148, doi: 10.1016/j.palaeo.2013.02.029.
- Guex, J., 1995, Ammonites hettangiennes de la Gabbs Valley Range, Nevada: Memoire Geologie Lausanne, v. 23, p. 1–130.
- Guex, J., Rakus, M., Taylor, D., and Bucher, H., 1997, The Triassic-Jurassic boundary: Proposal for stratotype defined in the Gabbs Valley Range (Nevada): International Subcommittee Jurassic Stratigraphy Newsletter, v. 24, p. 26–30.
- Guex, J., Bartolini, A., Atudorei, V., and Taylor, D., 2004, High-resolution ammonite and carbon isotope stratigraphy across the Triassic-Jurassic boundary at NYC (Nevada): Earth and Planetary Science Letters, v. 225, p. 29–41, doi: 10.1016/j.epsl.2004.06.006.
- Hallam, A., and Wignall, P.B., 2000, Facies changes across the Triassic-Jurassic boundary in Nevada, USA: Journal of the Geological Society, v. 157, p. 49–54, doi: 10.1144/jgs.157.1.49.
- Hautmann, M., Benton, M.J., and Tomas˘ových, A., 2008, Catastrophic ocean acidification at the Triassic-Jurassic boundary: Neues Jahrbuch für Geologie und Palaontologie Abhandlungen, v. 249, p. 119–127, doi: 10.1127/0077-7749/2008/0249-0119.
- Hoegh-Guldberg, O., Mumby, P.J., Hooten, A.J., Steneck, R.S., Greenfield, P., Gomez, E., Harvell, C.D., Sale, P.F., Edwards, A.J., Caldeira, K., Knowlton, N., Eakin, C.M., Iglesias-Prieto, R., Muthiga, N., Bradbury, R.H., Dubi, A., and Hatziolos, M.E., 2007, Coral reefs under rapid climate change and ocean acidification: Science, v. 318, p. 1737–1742, doi: 10.1126/science.1152509.
- Kiessling, W., Roniewicz, E., Villier, L., Léonide, P., and Struck, U., 2009, An early Hettangian coral reef in southern France: Implications for the end-Triassic reef crisis: Palaios, v. 24, p. 657–671, doi: 10.2110/palo.2009.p09-030r.
- Lathuilière, B., and Marchal, D., 2009, Extinction, survival and recovery of corals from the Triassic to Middle Jurassic time: Terra Nova, v. 21, p. 57–66, doi: 10.1111/j.1365-3121.2008.00856.x.
- Lucas, S.G., Taylor, D.G., Guex, J., Tanner, L.H., and Krainer, K., 2007, Updated Proposal for Global Stratotype Section and Point for the base of the Jurassic System in the NYC area, Nevada, USA: ISJJS Newsletter, v. 34, p. 34–42.
- Martindale, R.C., Berelson, W.M., Corsetti, F.A., Bottjer, D.J., and West, A.J., 2012, Constraining carbonate chemistry at a potential ocean acidification event (the Triassic-Jurassic boundary) using the presence of corals and coral reefs in the fossil record: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 350–352, p. 114–123, doi: 10.1016/j.palaeo.2012.06.020.
- McRoberts, C.A., Ward, P.D., and Hesselbo, S., 2007, A proposal for the base Hettangian Stage (base Jurassic System) GSSP at NYC (Nevada, USA) using carbon isotopes: International Subcommission on Jurassic Stratigraphy Newsletter, v. 34, p. 43–49.
- Melnikova, G.K., and Roniewicz, E., 2012, Early Jurassic corals of the Pamir Mountains—A new Triassic-Jurassic transitional fauna: Geologica Belgica, v. 15, p. 376–381.
- Muller, S.W., and Ferguson, H.G., 1936, Triassic and Lower Jurassic Formations of West Central Nevada: GSA Bulletin, v. 47, p. 241–252, doi: 10.1130/GSAB-47-241.
- Porter, S.J., Smith, P.W., Caruthers, A.H., Hou, P., Grocke, D.R., and Selby, D., 2014, New high-resolution geochemistry of Lower Jurassic marine sections in western North America: A global positive isotope excursion in the Sinemurian?: Earth and Planetary Science Letters, v. 397, p. 19–31, doi: 10.1016/j.epsl.2014.04.023.
- Prinz, P., 1991, Mezozoiche Korallen aus Nordchile: Palaeontographica, v. 216, p. 147–209.
- Ritterbush, K.A., Bottjer, D.J., Corsetti, F.A., and Rosas, S., 2014, New evidence on the role of siliceous sponges in ecology and sedimentary facies development in Eastern Panthalassa following the Triassic-Jurassic mass extinction: Palaios, v. 29, p. 652–668, doi: 10.2110/palo.2013.121.
- Roniewicz, E., and Michalík, J., 2002, Carnian Corals from the Male Karpaty Mountains, Western Carpathians, Slovakia: Geologica Carpathica, v. 53, p. 149–157.
- Roniewicz, E., and Morycowa, E., 1993, Evolution of the Scleractinia in the light of microstructural data: Courier Forschungsinstitut Senckenberg, v. 164, p. 233–240.
- Roniewicz, E., and Stanley, G.D., Jr., 2013, Upper Triassic corals from Nevada, western North America, and the implications for paleoecology and paleogeography: Journal of Paleontology, v. 87, no. 5, p. 934–964, doi: 10.1666/12-081.
- Senowbari-Daryan, B., and Stanley, G.D., Jr., 1994, Lower Jurassic marine carbonate deposits in central Peru: Stratigraphy and paleontology: Palaeontographica Abteilung A, v. 233, p. 43–56.
- Sha, J., 2002, Hispanic Corridor formed as early as Hettangian: On the basis of bivalve fossils: Chinese Science Bulletin, v. 47, p. 414–417, doi: 10.1360/02tb9096.
- Shepherd, H.M.E., 2013, Nearing the End: Reef Building Corals and Bivalves in the Late Triassic and Comparing Corals and Bivalves Before and After the End-Triassic Mass Extinction Using a Taxonomic Database [M.S. Thesis]: Missoula, the University of Montana, 90 p.
- Smith, P.L., 1983, The Pliensbachian ammonite Dayiceras dayiceroides and Early Jurassic paleogeography: Canadian Journal of Earth Sciences, v. 20, p. 86–91, doi: 10.1139/e83-008.
- Stanley, G.D., Jr., 1997, Evolution of reefs of the Mesozoic: Proceedings of the 8th International Coral Reef Symposium, p. 1657–1662.
- Stanley, G.D., Jr., 2003, The evolution of modern corals and their early history: Earth-Science Reviews, v. 60, no. 3, p. 195–225.
- Stanley, G.D., Jr., and Beauvais, L., 1994, Corals from an Early Jurassic coral reef in British Columbia: Refuge on an oceanic island reef: Lethaia, v. 27, p. 35–47, doi: 10.1111/j.1502-3931.1994.tb01553.x.
- Stanley, G.D., Jr., and McRoberts, C.A., 1993, A coral reef in the Telkwa Range, British Columbia: The earliest Jurassic example: Canadian Journal of Earth Sciences, v. 30, p. 819–831, doi: 10.1139/e93-068.
- Taylor, D.G., Smith, P.L., Laws, R.A., and Guex, J., 1983, The stratigraphy and biofacies trends of the Lower Mesozoic Gabbs and Sunrise formations, west-central Nevada: Canadian Journal of Earth Sciences, v. 20, p. 1598–1608, doi: 10.1139/e83-149.
- Taylor, D.G., Guex, J., and Rakus, M., 2001, Hettangian and Sinemurian Ammonoid Zonation for the Western Cordillera of North America: Bulletin de Géologie de l’ Université de Lausanne, v. 87, p. 381–421.
- Ward, P.D., Garrison, G.H., Williford, K.H., Kring, D.A., Goodwin, D., Beattie, M.J., and McRoberts, C.A., 2007, The organic carbon isotopic and paleontological record across the Triassic-Jurassic boundary at the candidate GSSP section at Ferguson Hill, Muller Canyon, Nevada, USA: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 244, p. 281–289, doi: 10.1016/j.palaeo.2006.06.042.
- Wells, J.W., 1953, Mesozoic invertebrate faunas of Peru, Lower Jurassic corals from the Arequipa region: Novitates of the American Museum, v. 1631, p. 1–14.
