Introduction
The Great Acceleration in human population growth and anthropogenic environmental modification in the
mid-twentieth century represents the beginning of a proposed Anthropocene Epoch. This was preceded at ca. 3.5 Ma
by a great acceleration in the rate of evolutionary brain growth (encephalization) in human ancestors (hominins)
that coincided with initiation of the earliest archaeological evidence for fabrication and use of stone tools
(McPherron et al., 2010; Harmand et al., 2015; Du et al., 2018). The association of encephalization with
stone-tool fabrication and use is consistent with the concept that the benefits of stone-tool use contributed to
evolutionary selective pressures for greater cognitive abilities. Increasing cognitive abilities and associated
creation of increasingly sophisticated tools led eventually, and perhaps inexorably, to the Anthropocene.
Early stone-tool fabrication and use marked the beginning of a suite of behavioral and anatomical changes
associated with encephalization, but the nature and timing of these changes and their relative significance for
encephalization are poorly understood. Increased cognitive abilities applied toward stone-tool use appear to
have led to additional behaviors and skills that conferred additional survival advantages. This article is an
attempt to evaluate the origins of stone-tool use and the significance of associated changes, both as causes and
consequences of early encephalization.
Hominin Brain Growth and Tool Use
The late Miocene African hominin Sahelanthropus tchadensis, from the Lake Chad region of central
Africa and dated at ca. 7 Ma, is the oldest known hominin with evidence for upright posture as indicated by the
morphology of a partial femur and a digitally reconstructed fossil skull (Daver et al., 2022). Endocranial
volume (inside of the skull), at 360–370 cm3, is similar to that of chimpanzees (Zollikofer et al.,
2005). Two partial skulls of the younger Pliocene hominins Ardipithecus ramidus and
Australopithecus anamensis have similar brain volume (Suwa et al., 2009; Haile-Selassie et al., 2019).
At ca. 3.5 Ma, the rate of brain growth accelerated dramatically from near zero to an average rate of
~4.2%/105 yr, reaching a modern volume of ~1350 cm3 that is almost quadruple its early
Pliocene value (Fig. 1; Du et al., 2018). Numerous skeletal features indicate that the transition to rapid
encephalization occurred after adaptation to upright posture, although some of these features have been
interpreted to indicate a mix of arboreal and terrestrial locomotion.
 Figure
1
Figure
1
Endocranial volume vs. fossil age for hominins over 0–7 Ma and the chronology of early hominin archaeological
sites. Endocranial volume data from compilation of Du et al. (2018), with additional points for the older
hominins
S. tchadensis (Zollikofer et al., 2005),
A. ramidus (Suwa et al., 2009), and
A.
anamensis (Haile-Selassie et al., 2019). Least-squares linear regression of data points <3.5 Ma
reveals an average volume increase of 4.2%/10
5 yr (~0.001% per generation; coefficient of
determination
r2 = 0.78). Green indicates background hominin endocranial volume before ca.
3.5 Ma acceleration and is projected forward to the present, where it encompasses the average brain volume of
modern chimpanzees. Note that various data points do not necessarily represent human ancestors and could be
now-extinct branches from the human ancestral lineage (e.g., Du et al., 2018; Diniz-Filho et al., 2019).
Estimated times of occasional, habitual, and obligatory stone-tool use are from Shea (2017a).
From its beginning at ca. 3.5 Ma, rapid encephalization was associated with stone-tool production and use in
eastern Africa. The subtle beginnings of tool use are represented by 3.4-Ma animal bones with scrapes and cut
marks interpreted as products of stone-tool butchery (McPherron et al., 2010) and by crude, 3.3-Ma stone tools
that were likely used for chopping, scraping, and/or crushing animal and/or plant matter (Harmand et al., 2015).
The oldest known stone tools, at Lomekwi 3 on the west side of Lake Turkana in northwestern Kenya (Fig. 2),
consist of locally available basalt and alkalic basaltic rocks. Compared to more abundant and younger stone
artifacts, the Lomekwi 3 cores are unusually massive and were broken with a low level of skill, perhaps by
block-on-block percussion (Fig. 3A; Harmand et al., 2015).
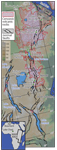 Figure 2
Figure 2
Relief map of the east African rift system, with rectangular symbols on the downthrown side of normal faults.
Also shown are the distribution of volcanic rocks associated with the rift system and areas where significant
numbers of hominin-related stone tools and/or bones have been recovered (green squares). These areas include all
22 stone-tool sites included in the compilation of stone-tool dimensions by Braun et al. (2019).
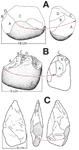 Figure
3
Figure
3
Examples of stone-tool artifacts. Red arrows and lettered faces show correlation between features with different
view direction. (A) Lomekwian (3.3 Ma) core from the west of Lake Turkana, Kenya (simplified from Harmand et
al., 2015). (B) Oldowan core (1.7 Ma) from Olduvai Gorge, Tanzania (redrawn from Leakey, 1971). (C) Middle to
late Pleistocene Acheulian core from Santa Ana Cave, Spain (simplified from García-Vadillo et al., 2022).
Oldowan Tools
Oldowan tools are simple stone tools produced at ca. 1.6–2.6 Ma by impact removal of rock chips from a “core”
to produce sharp but generally irregular edges. These tools are not as sophisticated as younger Acheulian stone
tools produced, for example, by coarse knapping to yield an approximately shaped tool followed by fine chip
removal to produce straight, or more evenly curved, sharp edges. (Attaching sharpened stones to wood to make
axes or stone-tipped spears occurred much later [<300 ka].) Oldowan tools appear to have been made by hitting
a stone held in one hand with a stone held in the other (pebble-core reduction), hitting or throwing a stone at
another stone or stone surface (anvil percussion), or placing the target stone on a stone surface and hitting
the target stone with another stone held in the other hand (bipolar-core reduction; e.g., Shea, 2017b). The rock
chips themselves may have been desired products. Tools made by these methods are not generally distinctive of
tool-making cultures or geographic areas and are simple enough that cultural transmission might not have been
necessary, leading to the inference of occasional rather than habitual fabrication (Fig. 1; Shea, 2017a).
A compilation of measurements of rocks interpreted as cores (targets of tool fabrication) by hominins at 21
eastern African archaeological sites, all dated to between 1.4 and 2.6 Ma, includes the average of the maximum
dimension of the cores from each site (Braun et al., 2019). The average of the averages from all 21 sites, 6.5 ±
3.7 cm (2σ) (equivalent to 2.5 ± 1.5 in.), indicates that these cores are generally small, consistent perhaps
with the small size and less developed thumbs of early hominins (Grabowski et al., 2015; Karakostis et al.,
2021). These small tools are inferred to have been used for butchery because cut and scrape marks on associated
animal bones have been found at some of the sites (e.g., de Heinzelin et al., 1999, Bouri, Afar, Ethiopia,
2.5 Ma; Domínguez-Rodrigo et al., 2005, Gona, Afar, Ethiopia, 2.1–2.6 Ma; Ferraro et al., 2013,
Kanjera South, Kenya, ca. 2.0 Ma). Stone tools from many of these sites include subangular to
subrounded pebbles and cobbles that are partially knapped (Figs. 3A–3B) so that a hominin could hold at least
some of the smoother side or end of the tool while chopping, sawing, or scraping with the sharp edge of the
other side or end (“pebble tools”; e.g., Leakey, 1971; Stout et al., 2010; Harmand et al., 2015; Shea, 2017b).
Some clasts may have been selected for their planar surfaces that would provide natural targets for percussive
removal of chips at the edges (Goldman-Neuman and Hovers, 2012).
Regardless of the general simplicity of Oldowan stone tools, some tools were crafted with significant skill.
Reassembly of stone tools and chipped debris at the 2.34 Ma Lokalalei site west of Lake Turkana in western Kenya
indicates skill and consistency in exploiting the edges and faces of cores during tool fabrication (Roche et
al., 1999). Reassembly and analysis of artifacts from three of the oldest known Oldowan tool sites (ca. 2.6 Ma)
at Gona in the Afar region of eastern Ethiopia indicates that cores “were efficiently reduced through the
production of large, invasive flakes, using a range of strategies comparable to that seen in later Oldowan
times” (Stout et al., 2010, 488). Two-handed pebble-core reduction with good manual dexterity was inferred, with
the suggestion of cultural transmission of knapping technique that influenced core-reduction strategies (Stout
et al., 2010).
Rock-Type Selectivity
Production of stone tools requires selection of rock types with appropriate mechanical properties, as most
rocks would not make good cutting or scraping tools and some are difficult to break. Stone tools were likely
intended for some purpose that the stone knapper had in mind, thus requiring forethought, although the
specificity of forethought might have been only of a generally sharp object with multiple potential uses.
A variety of hominin preferences can be discerned from stone artifact assemblages from different geologic
environments. An early example of rock selectivity is apparent for 2.5–2.6 Ma stone tools from sites in Gona,
Afar, in which the stone tools were derived from local conglomerate or stream-bed gravel containing a variety of
clast types. Hominins preferred aphanitic and vitreous volcanic rocks and avoided basalt (Stout et al., 2005).
At another archaeological site near the lower Awash River, ca. 2.6 Ma hominins preferred rhyolite, dacite, and
cryptocrystalline silica and avoided basalt (Braun et al., 2019). Durability of stone tools rather than fracture
predictability appears to have influenced rock selection at Kanjera South in western Kenya (Braun et al., 2009).
At Olduvai Gorge, late Oldowan and early Acheulian artifact assemblages (ca. 1.7–1.4 Ma) include large fractions
made of quartzite, which was apparently preferred because angular fragments from nearby bedrock outcrops
presented attractive faces and angles for flake removal, unlike the generally rounded shapes characteristic of
volcanic rocks forming streambed pebbles and cobbles (McHenry and de la Torre, 2018).
Tool Use
The uses of Oldowan tools for purposes other than butchery are not apparent from the tools themselves, but
perhaps can be discerned from inventories of vertebrate bones from areas of hominin butchery. At Kanjera South
in western Kenya (Fig. 2), three excavations of ca. 2.0 Ma fluvial and lacustrine strata near the shore of Lake
Victoria yielded several thousand bones, bone fragments, and lithic artifacts (Ferraro et al., 2013). An
inventory of the bones indicates that hominins butchered small herbivores generally as whole or nearly whole
carcasses, whereas medium-sized herbivore bones are disproportionately overrepresented by skulls and long limb
bones. This was interpreted to indicate that small herbivores were hunted and whole carcasses brought to sites
of butchery, while medium-sized herbivores were scavenged for parts that contain nutritious marrow and brain.
Demographic profiles of bovid bones at the 1.8 Ma FLK Zinj archaeological site at Olduvai Gorge, Tanzania (Fig.
2), approximate natural herd demographics rather than the greater abundance of the old and young that
characterize lion kills and scavenged remains (Bunn and Gurtov, 2014). Ambush hunting, most likely with wooden
spears sharpened with abrasive stones, was inferred for these Olduvai bones.
Hunting small- to medium-size mammals with wooden spears seems likely in eastern Africa by 2 Ma, regardless of
the lack of direct evidence for spear fabrication and use. Evidence of hominin wood use is indirect or
inferential as wood does not survive burial under generally warm, oxidizing soil conditions and so is absent in
fluvial, paleosol, and subaerially exposed lake-margin strata that contain African hominin fossil and bone
matter. Wooden spears are preserved in 300 ka anoxic lake sediments deposited under temperate to arctic
conditions in northern Europe (Schoch et al., 2015), but similar preservation is unknown at African hominin
fossil sites. Important points here are that ambush hunting with wooden spears would have led to more consistent
meat-rich diets and, with sufficient hunting success, habitual rather than occasional fabrication and use of
stone tools for carcass butchery.
Environmental Consequences
The appearance in eastern Africa of hominins armed with wooden spears and butchery tools, whenever it occurred,
might be expected to have had ecological consequences. Gradual drying and the spread of C4 grasses
since ca. 4 Ma in eastern Africa (Cerling et al., 2011) was associated with declining megaherbivore diversity
(Faith et al., 2018; Bibi and Cantalapiedra, 2023). These changes began, however, before the archaeological
appearance of the oldest known stone tools at 3.3 Ma and well before evidence of hominin hunting of small
herbivores at Kanjera South at ca. 2.0 Ma (Ferarro et al., 2013) and larger bovids at the 1.8 Ma FLK Zinj
archaeological site at Olduvai Gorge (Bunn and Gurtov, 2014).
The fossil record of carnivores, however, is different than that of herbivores. Statistical analysis of bone
and tooth remains from eastern Africa indicates that a major reduction in the diversity of large carnivores
(>21.5 kg) began at ca. 2.0 Ma (Werdelin and Lewis, 2013). Another analysis determined that extinction rates
of large carnivores began to increase at ca. 4 Ma and occurred at increasing rates up to the present (Faurby et
al., 2020). Smaller carnivores, however, did not experience increased extinction rates during this time
interval. Early extinction of large carnivores was attributed to loss of carcasses due to direct confrontation
with hominins, potentially wielding spears (kleptoparasitism), and to hominin scavenging of undefended carcasses
(Faurby et al., 2020). The increasing rate of extinction over time was attributed to progressive reduction of
available prey because of increasingly effective hunting and scavenging by hominins.
A New Diet and a New Body
A variety of anatomical and behavioral changes accompanied encephalization. The size of hominin teeth began
shrinking at ca. 2 Ma, roughly at the transition from Australopithecus to Homo. Using stone
tools to process animal and plant matter into smaller fragments, tenderize meat, and remove animal tissues that
are mechanically resistant to chewing could all contribute to evolution of smaller teeth and chewing muscles
(Zink and Lieberman, 2016). Dental reduction has been linked to cooking, which softens food, destroys pathogens
and toxins, and increases nutritional value (Gowlett and Wrangham, 2013). Evolution toward a less flared rib
cage indicates less space for a smaller gut, consistent with digestion of cooked rather than raw foods (Andrews
and Johnson, 2019). Combined with encephalization, dental reduction changed skull shape, leading eventually to a
short mouth and greater capacity to shape sounds with the tip of the tongue (Lieberman, 2011), which likely
facilitated language development. Changes in the hominin body that reflect a new and changing lifestyle include
a longer and stronger thumb more capable of tool fabrication and use (Karakostis et al., 2021) and greater
adaptation to walking and running.
Dispersal
Regardless of crude fabrication skills and the possibility of only occasional tool use, early stone-tool–using
hominins dispersed out of tropical and subtropical Africa into temperate environments. Stone tools at Ain
Boucherit near the town of Beni Fouda in northeastern Algeria, dated at ca. 2.4 Ma and located less than 100 km
from the Mediterranean Sea, are ~4500 km away from eastern African hominin fossil sites (Fig. 2; Sahnouni et
al., 2018). By 2.12 Ma, hominins had reached the Loess Plateau near Xi’an in central China (Zhu et al., 2018).
Hominins at this time had developed the capacity to live in and disperse through diverse environments that were
increasingly temperate at more northern latitudes. Use of stone tools for butchery, wooden spears for hunting
and defense, sharpened sticks for digging for roots and tubers, and fire for warmth and cooking would have
enabled dispersal and survival in diverse environments, but it is uncertain if any or all of these factors were
particularly relevant to long-distance dispersals.
Encephalization Without Tools?
The crude nature of Oldowan tools, and the minuscule improvement in fabrication in the first ~1.5 m.y. of tool
use, allows for the possibility that early encephalization was only marginally related to tool fabrication and
use. Another likely cause of early hominin encephalization is an increasingly complex social environment with
larger social-group populations, as indicated by positive correlations between brain size and social group size
in primates (Dunbar, 2009). In other words, it takes a lot of brain power to form and maintain long-term social
bonds that generally provide mutual benefits to all parties. In this context, evolutionary development of the
extensive and uniquely complex kinship relations of human societies (Chapais, 2017) provided survival benefits
and selective pressures that contributed to encephalization. More recent (post-Oldowan) and more sophisticated
stone-tool use might have contributed more effectively to selective pressures for greater cognition and brain
growth.
The First Geologists?
The east African rift system is a divergent plate boundary that is gradually accommodating tectonic breakup of
eastern Africa and its separation from the rest of the continent (e.g., Martin, 2023). Volcanic activity
associated with rifting produced generally basaltic lava flows and alkalic volcanic and shallow intrusive rocks,
and locally produced volcanic rocks with a wide range of silica content, as is characteristic of continental
rifts. Where these volcanic rocks were non-vesicular and fresh (unaltered by weathering, diagenesis, or
hydrothermal processes), and hard but not too tough, they were commonly the dominant material selected by
hominins for tool fabrication. Toughness is a mechanical property that can make a rock difficult to break, even
with a steel rock hammer and a strong human arm. In contrast, glassy rocks such as obsidian are not very tough
but yield sharp edges when broken and are hard enough to sustain a sharp edge during light to moderate use.
Other types of volcanic rock, quartzite, quartz (probably vein quartz), and amorphous silica such as chert have
also been used, as they are typically hard and can yield sharp edges upon impact, but are not so tough as to
greatly inhibit breakage by stone-wielding hominins.
If earliest hominin use of sharp-edged stone tools resulted from use of chipped debris generated when breaking
bones or nuts, then areas underlain by suitable rock units would be necessary to foster the behavioral
transition from breaking with stones to cutting or scraping with broken stones. The east African rift system
south of Ethiopia consists of western and eastern branches, with abundant volcanic rocks associated only with
the eastern branch (Fig. 2). Sediment accumulation and erosional incision occurred along both branches, but
major early hominin sites are located almost entirely within or near the eastern branch. This spatial
relationship suggests that early hominin tool use was associated specifically with areas of geologically young
volcanic rocks. Thus, even if it is a stretch to think of tool-fabricating, small-brain hominins as geologists,
such a hominin appears to have been a product of a specific geologic environment.
There are other reasons why hominin fossils and tools from the western branch of the east African rift system
might be largely unknown, including more rainfall and vegetation that would conceal fossil-bearing strata, and
more adverse political and societal environments for archaeological fieldwork. The western branch is also
characterized by deep grabens with abundant lakes, perhaps concealing evidence of hominin tool use, while rift
shoulders could have been sites of persistent erosion rather than erosion with occasional sediment accumulation
and preservation. This would contrast with the eastern branch, where volcanic activity was associated with more
complex and varied structural and topographic changes that led in many areas to exhumation of Pliocene and
Pleistocene, fossil- and tool-bearing strata (e.g., Quade et al., 2004). The fact remains, however, that Lomekwi
3 and most Oldowan stone tools were derived from volcanic and shallow intrusive rocks in an environment of
abundant and geologically young volcanic rocks associated with the eastern rift.
Conclusion
Hominin encephalization began at about the same time as stone-tool production and carcass butchery with sharp
stone tools. This is consistent with the concept that early encephalization was triggered by the survival
benefits of making and using stone tools. The advantages of stone-tool–assisted butchery, however, seem
inadequate as the primary driver of early encephalization. A variety of other changes were associated with
encephalization, but the relative significance and timing of these changes in producing evolutionary selective
pressures for greater cognitive abilities and fine motor skills are poorly known. These changes include (1) use
of stone-sharpened wooden spears for hunting and defense; (2) use of stone-sharpened sticks to dig for roots and
tubers; (3) use of fire for cooking; (4) development of a stronger and more dexterous thumb; (5) development of
language; and (6) development of increasingly complex social relationships with increasing social-group size.
Regardless of these uncertainties, stone tools appear to have been foundational in triggering 3.5 m.y. of rapid
evolutionary brain growth. I suggest that sharp rock fragments produced inadvertently by bone and/or nut
cracking at sites located on hard and brittle volcanic rocks within the east African rift system provided a
setting where hominins learned to use the sharp edges of broken rocks for butchery and then gradually
transitioned to deliberate fabrication of stone tools. Effective tool production was then based partially on
increasing recognition of the variable mechanical properties of rocks relevant to stone-tool fabrication and
knowledge of the color, texture, and distribution of useful rock types. Increasingly complex communication among
hominins would have led to expressions for relevant rock properties and distributions, eventually leading to
exchanges of knowledge crudely resembling discussions among geologists.
Acknowledgments
I thank Steve Kuhn, Jay Quade, and Andy Cohen for constructive comments on earlier drafts, and an anonymous
GSA Today reviewer for a thoughtful and constructive review.
References Cited
- Andrews, P., and Johnson, R.J., 2019, Evolutionary basis for the human diet: Consequences for human health:
Journal of Internal Medicine, v. 287, p. 226–237, https://doi.org/10.1111/joim.13011.
- Bibi, F., and Cantalapiedra, J.L., 2023, Plio-Pleistocene African megaherbivore losses associated with
community biomass restructuring: Science, v. 380, no. 6649, https://doi.org/10.1126/science.add8366.
- Braun, D.R., Plummer, T., Ferraro, J.V., Ditchfield, P., and Bishop, L.C., 2009, Raw material quality and
Oldowan hominin tool-stone preferences: Evidence from Kanjera South, Kenya: Journal of Archaeological Science,
v. 36, p. 1605–1614, https://doi.org/10.1016/j.jas.2009.03.025.
- Braun, D.R., et al., 2019, Earliest known Oldowan artifacts at >2.58 Ma from Ledi-Geraru, Ethiopia,
highlight early technological diversity: Proceedings of the National Academy of Sciences of the United States
of America, v. 116, no. 24, p. 11,712–11,717, https://doi.org/10.1073/pnas.1820177116.
- Bunn, H.T., and Gurtov, A.N., 2014, Prey mortality profiles indicate that Early Pleistocene Homo at
Olduvai was an ambush predator: Quaternary International, v. 322–323, p. 44–53,
https://doi.org/10.1016/j.quaint.2013.11.002.
- Cerling, T.E., Wynn, J.G., Andanje, S.A., Bird, M.I., Korir, D.K., Levin, N.E., Mace, W., Macharia, A.N.,
Quade, J., and Remien, C.H., 2011, Woody cover and hominin environments in the past 6 million years: Nature,
v. 476, p. 51–56, https://doi.org/10.1038/nature10306.
- Chapais, B., 2017, From chimpanzee society to human society, in Muller, M.N., Wrangham, R.W., and
Pilbeam, D.R., eds., Chimpanzees and Human Evolution: Cambridge, Massachusetts, The Belknap Press of Harvard
University Press, p. 427–463.
- Daver, G., Guy, F., Mackaye, H.T., Likius, A., Boisserie, J.-R., Moussa, A., Pallas, L., Vignaud, P., and
Clarisse, N.D., 2022, Postcranial evidence of late Miocene hominin bipedalism in Chad: Nature, v. 609,
https://doi.org/10.1038/s41586-022-04901-z.
- de Heinzelin, J., Clark, J.D., White, T., Hart, W., Renne, P., WoldeGabriel, G., Beyene, Y., and Vrba, E.,
1999, Environment and behavior of 2.5-million-year-old Bouri hominids: Science, v. 284, p. 625–629,
https://doi.org/10.1126/science.284.5414.625.
- Diniz-Filho, J.A.F., Jardim, L., Mondanaro, A., and Raia, P., 2019, Multiple components of phylogenetic
non-stationarity in the evolution of brain size in fossil hominins: Evolutionary Biology, v. 46, no. 1, p.
47–59, https://doi.org/10.1007/s11692-019-09471-z.
- Domínguez-Rodrigo, M., Pickering, T.R., Semaw, S., and Rogers, M.J., 2005, Cutmarked bones from Pliocene
archaeological sites at Gona, Afar, Ethiopia: Implications for the function for the world’s oldest stone
tools: Journal of Human Evolution, v. 48, p. 109–121, https://doi.org/10.1016/j.jhevol.2004.09.004.
- Du, A., Zipkin, A.M., Hatala, K.G., Renner, E., Baker, J.L., Bianchi, S., Bernal, K.H., and Wood, B.A.,
2018, Pattern and process in hominin brain size evolution are scale-dependent: Proceedings of the Royal
Society B, v. 285, https://doi.org/10.1098/rspb.2017.2738.
- Dunbar, R.I.M., 2009, The social brain hypothesis and its implications for social evolution: Annals of Human
Biology, v. 36, no. 5, p. 562–572, https://doi.org/10.1080/03014460902960289.
- Faith, J.T., Rowan, J., Du, A., and Koch, P.L., 2018, Plio-Pleistocene decline of African megaherbivores: No
evidence for ancient hominin impacts: Science, v. 362, no. 6417, p. 938–941,
https://doi.org/10.1126/science.aau2728.
- Faurby, S., Silvestro, D., Werdelin, L., and Antonelli, A., 2020, Brain expansion in early hominins predicts
carnivore extinctions in East Africa: Ecology Letters, v. 23, p. 537–544, https://doi.org/10.1111/ele.13451.
- Ferraro, J.V., et al., 2013, Earliest archaeological evidence of persistent hominin carnivory: PLoS One, v.
8, no. 4, https://doi.org/10.1371/journal.pone.0062174.
- García-Vadillo, F.-J., Canals-Salomó, A., Rodríguez-Álvarez, X.-P., and Carbonell-Roura, E., 2022, The large
flake Acheulean with spheroids from Santa Ana Cave (Cáceres, Spain): Journal of Archaeological Science,
Reports, v. 41, https://doi.org/10.1016/j.jasrep.2021.103265.
- Goldman-Neuman, T., and Hovers, E., 2012, Raw material selectivity in Late Pliocene Oldowan sites in the
Makaamitalu Basin, Hadar, Ethiopia: Journal of Human Evolution, v. 62, p. 353–366,
https://doi.org/10.1016/j.jhevol.2011.05.006.
- Gowlett, J., and Wrangham, R.W., 2013, Earliest fire in Africa: Towards the convergence of archaeological
evidence and the cooking hypothesis: Azania, v. 48, p. 5–30, https://doi.org/10.1080/0067270X.2012.756754.
- Grabowski, M., Hatala, K.G., Jungers, W.L., and Richmond, B.G., 2015, Body mass estimates of hominin fossils
and the evolution of human body size: Journal of Human Evolution, v. 85, p. 75–93,
https://doi.org/10.1016/j.jhevol.2015.05.005.
- Haile-Selassie, Y., Melillo, S.M., Vazzana, A., Benazzi, S., and Ryan, T.M., 2019, A 3.8-million-year-old
hominin cranium from Woranso-Mille, Ethiopia: Nature, v. 573, p. 214–219,
https://doi.org/10.1038/s41586-019-1513-8.
- Harmand, S., et al., 2015, 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya: Nature, v.
521, https://doi.org/10.1038/nature14464.
- Karakostis, F.A., Haeufle, D., Anastopoulou, I., Moraitis, K., Hotz, G., Tourloukis, V., and Harvati, K.,
2021, Biomechanics of the human thumb and the evolution of dexterity: Current Biology, v. 31,
https://doi.org/10.1016/j.cub.2020.12.041.
- Leakey, M.D., 1971, Olduvai Gorge, volume 3: Excavations in beds I and II, 1960–1963: Cambridge, UK,
Cambridge University Press, 272 p.
- Lieberman, D.E., 2011, The evolution of the human head: Cambridge, Massachusetts, Harvard University Press,
756 p.
- Martin, A.K., 2023, Opposite microplate rotations on the East African Rift: Similarity to double saloon door
tectonics: Journal of African Earth Sciences, v. 198, https://doi.org/10.1016/j.jafrearsci.2022.104803.
- McHenry, L.J., and de la Torre, I., 2018, Hominin raw material procurement in the Oldowan-Acheulean
transition at Olduvai Gorge: Journal of Human Evolution, v. 120, p. 378–401,
https://doi.org/10.1016/j.jhevol.2017.11.010.
- McPherron, S.P., Alemseged, Z., Marean, C.W., Wynn, J.G., Reed, D., Geraads, D., Bobe, R., and Béarat, H.A.,
2010, Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika,
Ethiopia: Nature, v. 466, p. 857–860, https://doi.org/10.1038/nature09248.
- Quade, J., Levin, N., Semaw, S., Stout, D., Renne, P., Rogers, M., and Simpson, S., 2004, Paleoenvironments
of the earliest stone toolmakers, Gona, Ethiopia: Geological Society of America Bulletin, v. 116, p.
1529–1544, https://doi.org/10.1130/B25358.1.
- Roche, H., Delagnes, A., Brugal, J.-P., Feibel, C., Kibunjia, M., Mourre, V., and Texier, P.-J., 1999, Early
hominid stone tool production and technical skill 2.34 Myr ago in West Turkana, Kenya: Nature, v. 399, p.
57–60, https://doi.org/10.1038/19959.
- Sahnouni, M., et al., 2018, 1.9-million- and 2.4-million-year-old artifacts and stone tool-cutmarked bones
from Ain Boucherit, Algeria: Science, v. 362, no. 6420, https://doi.org/10.1126/science.aau0008.
- Schoch, W.H., Bigga, G., Böhner, U., Richter, P., and Terberger, T., 2015, New insights on the wooden
weapons from the Paleolithic site of Schöningen: Journal of Human Evolution, v. 89, p. 214–225,
https://doi.org/10.1016/j.jhevol.2015.08.004.
- Shea, J.J., 2017a, Occasional, obligatory, and habitual stone tool use in hominin evolution: Evolutionary
Anthropology, v. 26, p. 200–217, https://doi.org/10.1002/evan.21547.
- Shea, J.J., 2017b, Stone Tools in Human Evolution: Behavioural Differences among Technological Primates: New
York, Cambridge University Press, 236 p.
- Stout, D., Quade, J., Semaw, S., Rogers, M.J., and Levin, N.E., 2005, Raw material selectivity of the
earliest stone toolmakers at Gona, Afar, Ethiopia: Journal of Human Evolution, v. 48, p. 365–380,
https://doi.org/10.1016/j.jhevol.2004.10.006.
- Stout, D., Semaw, S., Rogers, M.J., and Cauche, D., 2010, Technological variation in the earliest Oldowan
from Gona, Afar, Ethiopia: Journal of Human Evolution, v. 58, p. 474–491,
https://doi.org/10.1016/j.jhevol.2010.02.005.
- Suwa, G., Asfaw, B., Kono, R.T., Kubo, D., Lovejoy, C.O., and White, T.D., 2009, The Ardipithecus
ramidus skull and its implications for hominid origins: Science, v. 326,
https://doi.org/10.1126/science.1175825.
- Werdelin, L., and Lewis, M.E., 2013, Temporal change in functional richness and evenness in the eastern
African Plio-Pleistocene carnivoran guild: PLoS One, v. 8, no. 3,
https://doi.org/10.1371/journal.pone.0057944.
- Zhu, Z., et al., 2018, Hominin occupation of the Chinese Loess Plateau since about 2.1 million years ago:
Nature, v. 559, https://doi.org/10.1038/s41586-018-0299-4.
- Zink, K.D., and Lieberman, D.E., 2016, Impact of meat and Lower Palaeolithic food processing techniques on
chewing in humans: Nature, v. 531, p. 500–503, https://doi.org/10.1038/nature16990.
- Zollikofer, C.P.E., Ponce de León, M.S., Lieberman, D.E., Guy, F., Pilbeam, D., Likius, A., Mackaye, H.T.,
Vignaud, P., and Brunet, M., 2005, Virtual cranial reconstruction of Sahelanthropus tchadensis:
Nature, v. 434, p. 755–759, https://doi.org/10.1038/nature03397.