Introduction
The health and diversity of natural ecosystems—and human civilization—depend on our coordinated responses
to global changes that threaten earth’s long-term habitability. Soils, the thin veneer on the global
land surface that supports terrestrial life, are an integral component of anthropogenic climate change
mitigation strategies (Paustian et al., 2016; Loisel et al., 2019). Soils are a necessary part of the
solution for human-induced climate change because they represent one of the largest terrestrial carbon
(C) reservoirs, storing twice as much C as the earth’s atmosphere and vegetation combined (up to 2500 Pg
C; IPCC, 2013; Friedlingstein et al., 2020). Terrestrial C pools are a powerful C sink, with the
potential to offset up to 30% of anthropogenic C emissions, where some of the sequestered C persists in
soil over millennial time scales (Friedlingstein et al., 2020). Because of the relative sizes of the
different C reservoirs, even slight changes in the amount of C stored in soil can represent significant
changes in the global atmospheric concentration of carbon dioxide (CO2) and the earth’s
climate future.
How do we unlock soil’s potential for combating climate change? An important component of a comprehensive
response is to store more C in soils, particularly in soil pools that cycle C at slower rates compared
to the other reservoirs (ex., atmosphere, biomass, and on near surface soil layers) (Schmidt et al.,
2011). The amount of carbon stored in soil (soil organic C or SOC) is a balance between inputs and
outputs of carbon (Berhe, 2019a; Lavallee and Cotrufo, 2020). SOC storage in a given area (plot,
catchment, region, or another spatially constrained system) has been likened to a bank account, where
the “balance” is the bulk SOC stock or inventory (Fig. 1). Bank “deposits” are contributed by vegetation
litter, root exudates, living soil biota, deposition of eroded C, and remains of formerly living
organisms. The depletion of the balance in the soil carbon bank account is driven by microbial
decomposition of organic C inputs to CO2 and dissolved and particulate transport of C through
leaching and/or erosion.
 Figure
1
Figure
1
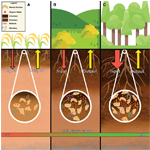
Soil organic carbon (SOC) is a dynamic and complex admixture. Here, three contrasting ecosystems reveal
differing SOC richness and dynamics: (A) agricultural, (B) grassland/shrubland, and (C) forested.
Conventional agriculture (A) often leads to lower carbon stocks, and overall, less carbon input to the
soil carbon pool. Grasslands (B) can harbor plants with deeper and more extensive root systems, medium
to high amounts of SOC stock, and greater carbon inputs to the SOC pool. Forests (C) can have the
deepest rooting system, a high amount of soil C stock, greatest density of mineral-associated C, and
high rate of input of C to soils. Overall, organo-mineral association(s) and SOC pool is a function of
the “balance” of C inputs and outputs in the soil organic carbon “bank account.”
The SOC that exists in soil can be subdivided into “slow-cycling” and “fast-cycling” pools akin to
checking and savings accounts (Lavallee and Cotrufo, 2020), respectively. Slow-cycling C is either
mineral-associated C that is found physically protected in soil aggregates or chemically bound to the
surfaces of reactive soil minerals; both mechanisms restrict decomposition and associated losses of SOC,
allowing it to persist in soil for decadal to millennial time scales (Schmidt et al., 2011; Hemingway et
al., 2019). In contrast, fast-cycling C is more readily degradable and prone to physical transport in
shorter time scales (Schmidt et al., 2011; Hemingway et al., 2019). Fast C cycling, which is akin to
funds in a checking account, is critical for maintenance of life in soil, because decomposition is the
main mechanism that recycles nutrients needed by organisms that call the soil home (Janzen, 2006). Even
small, but sustained, deposits into the soil C savings account over time allow for long-term buildup of
C in the slow-cycling pool with significant potential for climate change mitigation.
Increasing urgency for addressing the global climate emergency demands that we reduce the release of
greenhouse gasses from burning of fossil fuels, while finding appropriate alternatives to draw down some
atmospheric carbon through soil carbon sequestration and other means. As we seek these solutions, it is
important to remember that decomposition of organic matter (i.e., withdrawal of some of the balance from
the soil carbon checking account) is a critical ecosystem process because decay of organic residue
provides essential nutrients for plants and microbes in soil (Janzen, 2006). For this reason, we cannot
expect zero withdrawals from the soil carbon bank and must figure out how we can continue to “invest” in
soil C to maximize its input and retention in the soil, thus preventing fast release of C as greenhouse
gasses to the atmosphere. Maintenance of soil health through “smart” management practices has been
proven to simultaneously achieve SOC sequestration and provision of clean air, water, and a functional
habitat (Billings et al., 2021; Kopittke et al., 2022). Here, we explore prevailing issues with
conventional soil management, vulnerability of SOC to loss in a changing world, and strategies to
alleviate climate-change impacts on soil resources. In this framework, we identify strategies for soil C
sequestration and ways to prevent “overspending” in an uncertain future marked by changing climate and
increased demands to ensure food and nutritional security of the growing human population.
Carbon Losses Due to Conventional Soil Use and Degradation
An increasing human population and onset of the industrial age led to an increased demand for food,
energy, and water resources, and overall intensification of the agricultural sector. With intensive
agricultural practices came large-scale degradation of the global soil resource that included increased
rates of soil erosion (i.e., loss from working lands) that outpaced new soil production by 1–2 order(s)
of magnitude, largely resulting from deforestation to clear land for agriculture, conventional tillage
practices, and overgrazing (Lal, 2004; Montgomery, 2007). Conventional land management practices cause
physical disturbance of soils and have historically promoted enhanced agricultural yields, to the
detriment of SOC content, topsoil thickness, and overall soil health and structural stability (Phillips
et al., 1980; Reganold et al., 1987; Amundson et al., 2015). The systematic exploitation and
modification of undisturbed soils has led to the resulting agricultural soils being dubbed
“domesticated,” lacking hallmark resilience of their wild predecessors (Amundson et al., 2015). Soil
domestication for agriculture also presents broader, associated ecosystem issues, such as diminished
biodiversity from engineered crop community monocultures, introduction of chemical pesticides to hydro-
and pedospheres, and the delivery of vast quantities of esp. nitrogen and phosphorus fertilizers to
coastal margins. Conservation tillage and organic farming have been proposed as alternative approaches
that enhance soil health and to limit unsustainable soil “mining” and associated SOC overspending
(Montgomery, 2007). Estimates maintain that tillage management, when paired with cropping systems, can
sequester 0.03–0.11 Pg C yr-1 (Follett, 2001). Despite these promising advances, human
civilization and associated changes in land use and land cover led to the loss of 120 Pg C in the upper
~2 m of soils since humans adopted agriculture, with the fastest rate of loss occurring in the past 200
years (Sanderman et al., 2018).
Land Use/Land-Use Change (LULUC) practices such as conventional agriculture, deforestation, and wetland
conversion contribute 10%–14% of overall anthropogenic greenhouse gas emissions (Paustian et al., 2016).
The SOC pools impacted by LULUC have the potential to release massive amounts of C to the atmosphere,
making the preservation of these environments critical to protect soil C from loss both by reducing
future releases of C from soil to the atmosphere (avoided fluxes) and promoting drawdown of C that is
already in the atmosphere (sequestration of atmospheric CO2). Deforestation was historically
practiced to clear land for agriculture, but also continues to occur due to urban development, logging,
and an increase in wildfire frequency and intensity. These activities can destabilize SOC, releasing
slow-cycling C stored even in deeper soil layers (Drake et al., 2019). This also lowers ecosystem
functions that SOC can provide, such as water retention and nutrient cycling (Veldkamp et al., 2020).
Similarly, histosols (wetland soils, including peatlands with no underlying permafrost) can play a
critical role because they make up only 1% of soils globally, yet contain a larger proportion of SOC
(179 Pg C, or ~12% of SOC in the upper 100 cm globally: Brady and Weil, 2017). This SOC accumulation can
be attributed to a lower rate of decomposition of SOC due to waterlogging and resultant limitation in
availability of free oxygen for the heterotrophic soil microorganisms that can otherwise effectively
decompose organic matter. Histosols have historically been targets for drainage and conversion to
high-yielding agricultural lands (Holden et al., 2004). Draining of histosols, due to atmospheric
warming and/or anthropogenic practices, can lead to rapid decomposition of SOC release to the atmosphere
(Couwenberg et al., 2011). Overall, the soil system stores large amounts of carbon, but it has continued
to experience rapid degradation due to human actions. However, adoption of climate-smart land management
practices has a clear potential to reduce the atmospheric CO2 burden and increase the amount
of carbon stored in the soil carbon bank, with multiple benefits for improving ecosystem health and
human welfare.
Vulnerability of SOC to Loss with Uncertain Future
Climate is a primary factor driving the rate of decomposition of SOC (Brady and Weil, 2017). Global
climate change can accelerate SOC losses due to increasing global atmospheric temperature, altered
precipitation patterns, and other changes (Bellamy et al., 2005; Walker et al., 2018). Warming often
increases the rate of microbial decomposition of SOC and subsequent CO2 efflux to the
atmosphere (Lloyd and Taylor, 1994; Lehmeier et al., 2013; Min et al., 2019). The effects of increasing
temperature on SOC losses vary with molecular complexity of SOC and environmental conditions (e.g.,
water limitation, aggregation, mineral association) (Davidson and Janssens, 2006). Complex SOC, with
high activation energy, is more sensitive to temperature than simple SOC (Lehmeier et al., 2013; Lefèvre
et al., 2014). The temperature sensitivity of protected, slow-cycling C has been less studied (Karhu et
al., 2019), which necessitates future studies that explore the relationship between slow-cycling C and
its sensitivity to environmental changes. Contrary to the positive relationship between temperature and
SOC decomposition rate, increases in water availability can increase (Kaiser et al., 2015; Min et al.,
2020) or decrease SOC decomposition (Freeman et al., 2001), depending on the systems of interest.
Precipitation can also indirectly affect SOC storage by inducing soil erosion, changes in pore
connectivity, and altering ecosystem structure (Pimentel et al., 1995; Smith et al., 2017; Wu et al.,
2018). In eroding landscapes, lateral distribution of topsoil C and its deposition in lower-lying
landform positions (Berhe et al., 2018) causes mixing of the relatively fast-cycling C with slow-cycling
C in deep soil layers.
The response of carbon stored in soil to climate change and other perturbations varies depending on the
nature of the soils and the type of change to the system (Berhe, 2019b). Here, we highlight how SOC will
respond to climate change using three important areas of concern and uncertainty (e.g., gelisols,
paleosols, and deep soil).
Gelisols
Gelisols are soils of very cold climate conditions and store ~1000 Pg C in the upper 3 m of active and
underlying layers of permafrost soils (Tarnocai et al., 2009; Hugelius et al., 2014). Gelisols have
accumulated C because of climate-driven slow decomposition rates (Ping et al., 2015; Turetsky et al.,
2020). Warming in the northern hemisphere is predicted to release 12.2–112.6 Pg C by 2100, according to
Representative Concentration Pathway 4.5 and 8.5 warming scenarios (IPCC, 2013). This huge uncertainty
in the projected C release in the northern hemisphere is partly due to considerable variability in
hydrology, soil conditions, and vegetation (McGuire et al., 2009; Schuur and Abbott, 2011; Ping et al.,
2015). The rapid destabilization of polar and high-altitude environments, often referred to as the most
sensitive barometers of climate change, serves as a benchmark for understanding anthropogenic
modifications to the global climate system.
Paleosols
Paleosols are soils that developed in different environmental conditions when topsoil was transported
downhill and buried by alluvial, colluvial, aeolian deposition, volcanic eruption, or human activities
over centuries to millennia (Marin-Spiotta et al., 2014; Chaopricha and Marin-Spiotta, 2014). This
process promotes SOC-mineral association(s) (Rumpel and Kögel-Knabner, 2011) that build up soil C stock
in the slow-cycling soil C savings account (Schmidt et al., 2011). Recent estimates suggest that
paleosol C is a significant global C reservoir (Lehmkuhl et al., 2016), but it is spatially variable
depending on landscape and climate history, thus making it difficult to estimate the total storage. The
effect of any environmental change on buried SOC is complex and poorly understood because paleosols are
not considered for the global C stock inventory and models. The possibility of the vast storage of SOC
raises questions on how the previously buried SOC will interact in the presence of water, modern soil
surface microbes, and addition of new fresh SOC, and finally if they will become a sink or a source of
greenhouse gasses in the presence of all the optimal conditions for decomposition.
Deep Soil
The overwhelming majority of soil C studies have focused on shallow soil depths, with little attention
paid to the amount of C stored in or the vulnerability of C in deep soil layers. Soils can develop to
>10 m depth, and deep soils (below 30 cm) can store up to 74% of the total profile C with radiocarbon
ages of 5,000–20,000 years old (Moreland et al., 2021). It is estimated that 28 Pg C is stored in soils
with deep weathered bedrock, suggesting that deep soil C is a large C reservoir that may be potentially
vulnerable to a changing climate (Moreland et al., 2021). Some soils are already showing evidence of
warming by 2 °C, since 1961, which has been observed at up to 3 m depths (Zhang et al., 2016). Although
decomposition rates are slower in deeper soils than in surface soils, recent studies have shown that
deep SOC is more vulnerable to loss than previously thought (Rumpel and Kögel-Knabner, 2011; Hicks Pries
et al., 2017; Min et al., 2020). Experimental warming to a depth of 1 m found that warming increased
annual soil respiration by ~35% and estimated that with a 4 °C increase, deep soils have the potential
to release 3.1 Pg C yr-1, equivalent to 30% of fossil fuel emissions (Hicks Pries et al.,
2017; Friedlingstein et al., 2020).
In the following section, we focus on “working lands,” where the global soil degradation problem can be
effectively addressed (in a cost- and time-efficient manner) through a suite of natural climate change
solutions.
Soils as Natural Climate Change Solutions
Intergovernmental Panel on Climate Change (IPCC) assessment reports and the Paris Agreement have
highlighted the importance of immediate action to prevent catastrophic changes to the earth system.
Inclusion of soils in local to global climate change mitigation strategies is a proven and
cost-effective strategy. Natural climate solutions can provide 37% of cost-effective CO2
mitigation necessary for a >66% chance of holding warming below 2 °C by 2030 (Griscom et al., 2017).
The “4 per 1000” effort has proposed soil as a natural climate change solution and endeavors to increase
SOC storage by 0.4% annually (Rumpel et al., 2020), thereby offsetting one third of global fossil fuel
emissions. Here, we provide a review of the available solutions to increase the amount of C stored in
the soil C savings account through a variety of land stewardship practices, including use of amendments
such as compost, biochar, waste, and management interventions such as reforestation, inclusion of deep
root perennials, and cover crops.
Restoring degraded lands and avoiding further land conversion (e.g., afforestation) can also help
mitigate climate change (Fig. 2; Table 1). Afforestation of degraded sites in the United States is
estimated to potentially sequester 2.43 Pg C yr-1 in the upper 30 cm of soil over 30 years
(Cook-Patton et al., 2020). Although afforestation efforts can increase SOC storage on decadal time
scales, the effects are largely site-specific. For example, depending on the prevailing climate of an
area, restoring grasslands might be a better option for C sequestration than
afforestation/reforestation, and converting grasslands to forest may yield less net SOC storage than
converting cropland to forest (Li et al., 2012; Bárcena et al., 2014). Soil restoration, specifically
for wetlands, has the potential to return these environments to a net C sink (Table 1; Waddington et
al., 2010) and represents a cost-efficient mitigation strategy—projected to cost ~US$20 per Mg of
sequestered C (Humpenöder et al., 2020).
 Figure
2
Figure
2
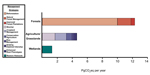
Various management strategies in forested, agriculture/grassland, and wetland ecosystems exhibit
differing propensities to take up CO
2. Overall, these strategies represent a way to expand
terrestrial ecosystem uptake of carbon (Friedlingstein et al., 2020; Paustian et al., 2016; Griscom et
al., 2017).
Regenerative agriculture (RA) also holds a substantial role in attaining negative carbon emissions from
rangeland and agricultural soils (Fig. 2; Table 1). RA is a set of locally adapted land practices that
minimize soil disturbance (e.g., no-till, minimum tillage, cover cropping) and losses (e.g., erosion,
degradation), while self-sustaining its ecosystem services (e.g., productivity, biodiversity;
Gonzalez-Sanchez et al., 2015) using agroecology-based theory and management (e.g., compost application,
crop, and grazing rotation, etc.). Hence, RA promotes C sequestration and soil health while
simultaneously reducing net SOC losses by providing a direct layer of protection from disturbance.
Ultimately, avoiding land conversion and disturbance a priori is the most effective strategy to maintain
SOC storage, as restoration of degraded lands accrues SOC slowly (Guo and Gifford, 2002). Both active
and preventative restoration practices are vital in providing ecosystem service co-benefits such as
water filtration and storage.
Land managers have added organic amendments to their soils since the early periods of agriculture. The
addition of C-rich amendments can improve soil health via enhancing nutrients and water storage, plant
productivity, microbial diversity, and soil structure (Woolf et al., 2010; Farooqi et al., 2018; Amelung
et al., 2020). Studies have now documented significant, positive impacts of organic amendments that
include a 2.3 Mg C ha-1 yr-1 increase in SOC stock in corn fields after six years
of biochar amendments (Blanco-Canqui et al., 2020), and a projected SOC sequestration potential of 1.2
Mg C ha-1 yr-1 in croplands after application of manure, sewage sludge, or straw
(Smith, 2004). In parts of the world that have large amounts of excess biomass (ex., agricultural
residue, manure, forest clippings, etc.), these amendments are viable options for climate change
mitigation (Fig. 2; Table 1), while at the same time replenishing C and nutrient stocks to increase the
ecosystem’s overall health and resilience (Koide et al., 2015).
Recent advances in plant-based strategies have also provided new insights to address net SOC loss. These
strategies rely on the ability of plants to self-regulate and self-optimize resource uptake and
allocation, and thus are considered cost-effective and sustainable with limited environmental
footprints. Plant roots are known to be a main source of SOC (Rasse et al., 2005), and root-derived SOC
is preferentially retained by minerals (Bird et al., 2008). Therefore, the introduction of roots into
deep soils can enhance slow-cycling C formation (Kell, 2011; Paustian et al., 2016). However, root
exudates enhance soil microbial activity and reduce SOC stock via priming (Fontaine et al., 2007;
Keiluweit et al., 2015). For this reason, plant roots are considered as a double-edged sword for SOC
formation (Dijkstra et al., 2021). Still, there is evidence that deeply rooting vegetation (esp.
perennial grasses) can sequester C into the deep soil (Slessarev et al., 2020). Extensive root systems
introduce C to the subsoil, enhancing SOC-mineral associations, aggregate protection, and reduced access
to SOC by soil microbes. In this manner, rhizosphere engineering benefits overall soil health and
resource use efficiency (Dessaux et al., 2016). With proper implementation, plant-based strategies can
synergize with existing strategies (e.g., conservation agriculture) to promote more SOC in the long-term
savings account (Fig. 2).
Concluding Thoughts
Soils have supported life and stored C throughout geological history. However, human civilization has
spurred drastic land use changes through agriculture and other activities. Additionally, profound
alteration to the global climate system has resulted from widespread fossil fuel utilization and
resulting greenhouse gas emissions. As we apply sophisticated models and propose novel technologies for
understanding and addressing anthropogenic climate change, a piece of the solution is found in the soil.
Natural climate change solutions involving soil health are not only cost effective, but also
non-negotiable, because they are key for securing the food, fuel, and fiber necessary for an
ever-increasing human population. Earth scientists, land managers, and policy makers must collaborate to
continue “spending” SOC while “investing” in SOC to increase its retention in the soil and maximize its
ability to support life. It’s a win-win climate solution that’s right beneath our feet. Let’s keep it
there.
Acknowledgments
This article was supported by the National Science Foundation (EAR 1623812) and University of California
Merced and Falasco Endowed Chair to AAB; and University of California Merced Chancellor’s Postdoctoral
Fellowship and the National Research Foundation of Korea (MSIT, NRF-2018R1A5A7025409) to KM. Icons used
in Figure 1 made by Freepik and Flat Icons, from www.flaticon.com.
References Cited
- Amelung, W., and 19 others, 2020, Towards a global-scale soil climate mitigation strategy: Nature
Communications, v. 11, 5427, https://doi.org/10.1038/s41467-020-18887-7.
- Amundson, R., Berhe, A.A., Hopmans, J.W., Olson, C., Sztein, A.E., and Sparks, D.L., 2015, Soil and
human security in the 21st century: Science, v. 348, https://doi.org/10.1126/science.1261071.
- Bárcena, T.G., Kiær, L.P., Vesterdal, L., Stefánsdóttir, H.M., Gundersen, P., and Sigurdsson, B.D.,
2014, Soil carbon stock change following af-forestation in Northern Europe: A meta-analysis: Global
Change Biology, v. 20, p. 2393–2405, https://doi.org/10.1111/gcb.12576.
- Bellamy, P.H., Loveland, P.J., Bradley, R.I., Lark, R.M., and Kirk, G.J.D., 2005, Carbon losses from
all soils across England and Wales 1978–2003: Nature, v. 437, p. 245–248,
https://doi.org/10.1038/nature04038.
- Berhe, A.A., 2019a, A climate change solution that’s right under our feet: A TEDtalk: TED, April
2019,
https://www.ted.com/talks/asmeret_asefaw_berhe_a_climate_change_solution_that_s_right_under_our_feet?language=en
(accessed 7 Feb. 2022).
- Berhe, A., 2019b, Drivers of soil change, in Busse, M., Giardina, C., Morris, D., and
Page-Dumroese, D., eds., Developments in Soil Science: Amsterdam, Elsevier,
https://doi.org/10.1016/B978-0-444-63998-1.00003-3.
- Berhe, A.A., Barnes, R.T., Six, J., and Marin-Spiotta, E., 2018, Role of soil erosion in
biogeochemical cycling of essential elements: Carbon, nitrogen, and phosphorous: Annual Review of
Earth and Planetary Sciences, v. 46, p. 521–548,
https://doi.org/10.1146/annurev-earth-082517-010018.
- Billings, S.A., and 17 others, 2021, Soil organic carbon is not just for soil scientists:
Measurement recommendations for diverse practitioners: Ecological Applications, v. 31, p. 1–19,
https://doi.org/10.1002/eap.2290.
- Bird, J., Kleber, M., and Torn, M.S., 2008, 13C and 15N stabilization dynamics in soil organic
matter fractions during needle and fine root decomposition: Organic Geochemistry, v. 39, p. 465–477,
https://doi.org/10.1016/j.orggeochem.2007.12.003.
- Blanco-Canqui, H., Laird, D.A., Heaton, E.A., Rathke, S., and Acharya, B.S., 2020, Soil carbon
increased by twice the amount of biochar carbon applied after 6 years: Field evidence of negative
priming: Global Change Biology. Bioenergy, v. 12, p. 240–251, https://doi.org/10.1111/gcbb.12665.
- Brady, N.C., and Weil, R., 2017, The nature and properties of soils: Columbus, Ohio, Pearson
Education Inc., 1104 p.
- Chaopricha, N., and Marin-Spiotta, E., 2014, Soil burial contributes to deep soil organic carbon
storage: Soil Biology & Biochemistry, v. 69, p. 251–264,
https://doi.org/10.1016/j.soilbio.2013.11.011.
- Cook-Patton, S., and 30 others, 2020, Mapping carbon accumulation potential from global natural
forest regrowth: Nature, v. 585, p. 545–550, https://doi.org/10.1038/s41586-020-2686-x.
- Couwenberg, J., Thiele, A., Tanneberger, F., Augustin, J., Bärisch, S., Dubovik, D., Liashchynskaya,
N., Michaelis, D., Minke, M., Skuratovich, A., and Joosten, H., 2011, Assessing greenhouse gas
emissions from peatlands using vegetation as a proxy: Hydrobiologia, v. 674, p. 67–89,
https://doi.org/10.1007/s10750-011-0729-x.
- Davidson, E.A., and Janssens, I.A., 2006, Temperature sensitivity of soil carbon decomposition and
feedbacks to climate change: Nature, v. 440, p. 165–173, https://doi.org/10.1038/nature04514.
- Dessaux, Y., Grandclément, C., and Faure, D., 2016, Engineering the rhizosphere: Trends in Plant
Science, v. 21, p. 266–278, https://doi.org/10.1016/j.tplants.2016.01.002.
- Dijkstra, F., Zhu, B., and Cheng, W., 2021, Root effects on soil organic carbon: A double-edged
sword: The New Phytologist, v. 230, p. 60–65, https://doi.org/10.1111/nph.17082.
- Drake, T.W., Van Oost, K., Barthel, M., Bauters, M., Hoyt, A.M., Podgorski, D.C., Six, J., Boeckx,
P., Trumbore, S.E., Cizungu Ntaboba, L., and Spencer, R.G.M., 2019, Mobilization of aged and
biolabile soil carbon by tropical deforestation: Nature Geoscience, v. 12, p. 541–546,
https://doi.org/10.1038/s41561-019-0384-9.
- Farooqi, Z., Sabir, M., Zeeshan, N., Naveed, K., and Hussain, M., 2018, Enhancing carbon
sequestration using organic amendments and agricultural practices, in Agarwal, R.K., ed.,
Carbon Capture, Utilization, and Sequestration: London, IntechOpen Limited,
https://doi.org/10.5772/intechopen.79336.
- Follett, R.F., 2001, Soil management concepts and carbon sequestration in cropland soils: Soil &
Tillage Research, v. 61, p. 77–92, https://doi.org/10.1016/S0167-1987(01)00180-5.
- Fontaine, S., Barot, S., Barré, P., Bdioui, N., Mary, B., and Rumpel, C., 2007, Stability of organic
carbon in deep soil layers controlled by fresh carbon supply: Nature, v. 450, p. 277–280,
https://doi.org/10.1038/nature06275.
- Freeman, C., Ostle, N., Kang, H., Lee, D.S., and Lee, J.S., 2001, An enzymic ‘latch’ on a global
carbon store: Nature, v. 409, p. 149–150, https://doi.org/10.1038/35051650.
- Friedlingstein, P., and 85 others, 2020, Global Carbon Budget 2020: Earth System Science Data, v.
12, p. 3269–3340, https://doi.org/10.5194/essd-12-3269-2020.
- Gonzalez-Sanchez, E.J., Veroz-Gonzalez, O., Blanco-Roldan, G.L., Marquez-Garcia, F., and
Carbonell-Bojollo, R., 2015, A renewed view of con-servation agriculture and its evolution over the
last decade in Spain: Soil & Tillage Research, v. 146, p. 204–212,
https://doi.org/10.1016/j.still.2014.10.016.
- Griscom, B.W., and 31 others, 2017, Natural climate solutions: Proceedings of the National Academy
of Sciences of the United States of America, v. 114, p. 11,645–11,650,
https://doi.org/10.1073/pnas.1710465114.
- Guo, L.B., and Gifford, R.M., 2002, Soil carbon stocks and land use change: A meta-analysis: Global
Change Biology, v. 8, p. 345–360, https://doi.org/10.1046/j.1354-1013.2002.00486.x.
- Hemingway, J.D., Rothman, D.H., Grant, K.E., Rosengard, S.Z., Eglinton, T.I., Derry, L.A., and Galy,
V.V., 2019, Mineral protection regulates long-term global preservation of natural organic carbon:
Nature, v. 570, p. 228–231, https://doi.org/10.1038/s41586-019-1280-6.
- Hicks Pries, C.E., Castanha, C., Porras, R.C., and Torn, M.S., 2017, The whole-soil carbon flux in
response to warming: Science, v. 355, p. 1420–1423, https://doi.org/10.1126/science.aal1319.
- Holden, J., Chapman, P.J., and Labadz, J.C., 2004, Artificial drainage of peatlands: Hydrological
and hydrochemical process and wetland restoration: Progress in Physical Geography, v. 28, p. 95–123,
https://doi.org/10.1191/0309133304pp403ra.
- Hugelius, G., and 16 others, 2014, Estimated stocks of circumpolar permafrost carbon with quantified
uncertainty ranges and identified data gaps: Biogeosciences, v. 11, p. 6573–6593,
https://doi.org/10.5194/bg-11-6573-2014.
- Humpenöder, F., Karstens, K., Lotze-Campen, H., Leifeld, J., Menichetti, L., Barthelmes, A., and
Popp, A., 2020, Peatland protection and restoration are key for climate change mitigation:
Environmental Research Letters, v. 15, p. 104093, https://doi.org/10.1088/1748-9326/abae2a.
- IPCC, 2013, Climate Change 2013: The Physical Science Basis, in Stocker, T.F., Qin, D.,
Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., and Midgley,
P.M., eds., Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental
Panel on Climate Change: Cambridge University Press, Cambridge, UK, and New York, 1535 p.
- Janzen, H.H., 2006, The soil carbon dilemma: Shall we hoard it or use it?: Soil Biology &
Biochemistry, v. 38, p. 419–424, https://doi.org/10.1016/j.soilbio.2005.10.008.
- Kaiser, M., Kleber, M., and Asefaw, A., 2015, How air-drying and rewetting modify soil organic
matter characteristics: An assessment to improve data interpretation and inference: Soil Biology
& Biochemistry, v. 80, p. 324–340, https://doi.org/10.1016/j.soilbio.2014.10.018.
- Karhu, K., and 11 others, 2019, Similar temperature sensitivity of soil mineral-associated organic
carbon regardless of age: Soil Biology & Biochemistry, v. 136, 107527,
https://doi.org/10.1016/j.soilbio.2019.107527.
- Keiluweit, M., Bougoure, J., Nico, P., Pett-Ridge, J., Weber, P., and Kleber, M., 2015, Mineral
protection of soil carbon counteracted by root exudates: Nature Climate Change, v. 5, p. 588–595,
https://doi.org/10.1038/nclimate2580.
- Kell, D., 2011, Breeding crop plants with deep roots: Their role in sustainable carbon, nutrient and
water sequestration: Annals of Botany, v. 108, p. 407–418, https://doi.org/10.1093/aob/mcr175.
- Koide, R.T., Nguyen, B.T., Skinner, R.H., Dell, C.J., Peoples, M.S., Adler, P.R., and Drohan, P.J.,
2015, Biochar amendment of soil improves resilience to climate change: Global Change Biology.
Bioenergy, v. 7, p. 1084–1091, https://doi.org/10.1111/gcbb.12191.
- Kopittke, P., et al., 2022, Ensuring planetary survival: The centrality of organic carbon in
balancing the multifunctional nature of soils: Critical Reviews in Environmental Science and
Technology, https://doi.org/10.1080/10643389.2021.2024484.
- Lal, R., 2004, Soil carbon sequestration impacts on global climate change and food security:
Science, v. 304, p. 1623–1627, https://doi.org/10.1126/science.1097396.
- Lavallee, J., and Cotrufo, F., 2020, Soil carbon is a valuable resource, but all soil carbon is not
created equal: The Conversation,
https://theconversation.com/soil-carbon-is-a-valuable-resource-but-all-soil-carbon-is-not-created-equal-129175
(accessed 7 Feb. 2022).
- Lefèvre, R., Barré, P., Moyano, F., Christensen, B., Bardoux, G., Eglin, T., Girardin, C., Houot,
S., Kätterer, T., van Oort, F., and Chenu, C., 2014, Higher temperature sensitivity for stable than
for labile soil organic carbon—Evidence from incubations of long-term bare fallow soils: Global
Change Biology, v. 20, p. 633–640, https://doi.org/10.1111/gcb.12402.
- Lehmeier, C.A., Min, K., Niehues, N.D., Ballantyne, I.V.F., and Billings, S.A., 2013,
Temperature-mediated changes of exoenzyme-substrate reaction rates and their consequences for the
carbon to nitrogen flow ratio of liberated resources: Soil Biology & Biochemistry, v. 57, p.
374–382, https://doi.org/10.1016/j.soilbio.2012.10.030.
- Lehmkuhl, F., Zens, J., Krauß, L., Schulte, P., and Kels, H., 2016, Loess-paleosol sequences at the
northern European loess belt in Germany: Distribution, geomorphology and stratigraphy: Quaternary
Science Reviews, v. 153, p. 11–30, https://doi.org/10.1016/j.quascirev.2016.10.008.
- Li, D., Niu, S., and Luo, Y., 2012, Global patterns of the dynamics of soil carbon and nitrogen
stocks following afforestation: A meta-analysis: The New Phytologist, v. 195, p. 172–181,
https://doi.org/10.1111/j.1469-8137.2012.04150.x.
- Lloyd, J., and Taylor, J.A., 1994, On the temperature dependence of soil respiration: Functional
Ecology, v. 8, p. 315–323, https://doi.org/10.2307/2389824.
- Loisel, J., Connors, J., Hugelius, G., Harden, J.W., and Morgan, C.L., 2019, Soils can help mitigate
CO2 emissions, despite the challenges: Proceedings of the National Academy of Sciences of
the United States of America, v. 116, p. 10,211–10,212, https://doi.org/10.1073/pnas.1900444116.
- Marin-Spiotta, E., Chaopricha, N.T., Plante, A.F., Diefendorf, A.F., Mueller, C.W., Grandy, A.S.,
and Mason, J.A., 2014, Long-term stabilization of deep soil carbon by fire and burial during early
Holocene climate change: Nature Geoscience, v. 7, p. 428–432, https://doi.org/10.1038/ngeo2169.
- McGuire, A.D., Anderson, L.G., Christensen, T.R., Scott, D., Laodong, G., Hayes, D.J., Martin, H.,
Lorenson, T.D., Macdonald, R.W., and Nigel, R., 2009, Sensitivity of the carbon cycle in the Arctic
to climate change: Ecological Monographs, v. 79, p. 523–555, https://doi.org/10.1890/08-2025.1.
- Min, K., Bagchi, S., Buckeridge, K., Billings, S.A., Ziegler, S.E., and Edwards, K.A., 2019,
Temperature sensitivity of biomass-specific microbial exo-enzyme activities and CO2
efflux is resistant to change across short- and long-term timescales: Global Change Biology, v. 25,
p. 1793–1807, https://doi.org/10.1111/gcb.14605.
- Min, K., Berhe, A.A., Khoi, C.M., van Asperen, H., Gillabel, J., and Six, J., 2020, Differential
effects of wetting and drying on soil CO2 concentration and flux in near-surface vs. deep
soil layers: Biogeochemistry, v. 148, p. 255–269, https://doi.org/10.1007/s10533-020-00658-7.
- Montgomery, D., 2007, Soil erosion and agricultural sustainability: Proceedings of the National
Academy of Sciences of the United States of America, v. 104, p. 13,268–13,272,
https://doi.org/10.1073/pnas.0611508104.
- Moreland, K., Tian, Z., Berhe, A., McFarlane, K., Hartsough, P., Hart, S., Bales, R., and O’Geen,
T., 2021, Deep in the Sierra Nevada critical zone: Saprock represents a large terrestrial organic
carbon stock: Environmental Research Letters, v. 16, p. 124059,
https://doi.org/10.1088/1748-9326/ac3bfe.
- Paustian, K., Lehmann, J., Ogle, S., Reay, D., Robertson, G.P., and Smith, P., 2016, Climate-smart
soils: Nature, v. 532, p. 49–57, https://doi.org/10.1038/nature17174.
- Phillips, R., Blevins, R., Thomas, G., Frye, W., and Phillips, S., 1980, No-tillage agriculture:
Science, v. 208, p. 1108–1113, https://doi.org/10.1126/science.208.4448.1108.
- Pimentel, D., Harvey, C., Resosudarmo, P., Sinclair, K., Kurz, D., McNair, M., Crist, S., Shpritz,
L., Fitton, L., Saffouri, R., and Blair, R., 1995, Environmental and economic costs of soil erosion
and conservation benefits: Science, v. 267, p. 1117–1123,
https://doi.org/10.1126/science.267.5201.1117.
- Ping, C., Jastrow, J., Jorgenson, M., Michaelson, G., and Shur, Y., 2015, Permafrost soils and
carbon cycling: Soil (Göttingen), v. 1, p. 147–171, https://doi.org/10.5194/soil-1-147-2015.
- Rasse, D.P., Rumpel, C., and Dignac, M., 2005, Is soil carbon mostly root carbon?: Plant and Soil,
v. 269, p. 341–356, https://doi.org/10.1007/s11104-004-0907-y.
- Reganold, J.P., Elliott, L.F., and Unger, Y.L., 1987, Long-term effects of organic and conventional
farming on soil erosion: Nature, v. 330, p. 370–372, https://doi.org/10.1038/330370a0.
- Rumpel, C., and Kögel-Knabner, I., 2011, Deep soil organic matter—A key but poorly understood
component of terrestrial C cycle: Plant and Soil, v. 338, p. 143–158,
https://doi.org/10.1007/s11104-010-0391-5.
- Rumpel, C., and 13 others, 2020, The 4p1000 initiative: Opportunities, limitations and challenges
for implementing soil organic carbon sequestration as a sustainable development strategy: Ambio, v.
49, p. 350–360, https://doi.org/10.1007/s13280-019-01165-2.
- Sanderman, J., Hengl, T., and Fiske, G.J., 2018, Soil carbon debt of 12,000 years of human land use:
Proceedings of the National Academy of Sciences of the United States of America, v. 115, p. E1700,
https://doi.org/10.1073/pnas.1706103114.
- Schmidt, M.W.I., and 13 others, 2011, Persistence of soil organic matter as an ecosystem property:
Nature, v. 478, p. 49–56, https://doi.org/10.1038/nature10386.
- Schuur, E.A.G., and Abbott, B., 2011, Climate change: High risk of permafrost thaw: Nature, v. 480,
p. 32–33, https://doi.org/10.1038/480032a.
- Siry, J.P., Cubbadge, F.W., and Ahmed, M.R., 2005, Sustainable forest management: Global trends and
opportunities: Forest Policy and Economics, v. 7, p. 551–561,
https://doi.org/10.1016/j.forpol.2003.09.003.
- Slessarev, E.W., Nuccio, E.E., McFarlane, K.J., Ramon, C.E., Saha, M., Firestone, M.K., and
Pett-Ridge, J., 2020, Quantifying the effects of switchgrass (Panicum virgatum) on deep
organic C stocks using natural abundance 14C in three marginal soils: Global Change Biology.
Bioen-ergy, v. 12, p. 1–14, https://onlinelibrary.wiley.com/doi/10.1111/gcbb.12729.
- Smith, A., Bond-Lamberty, B., Benscoter, B., Tfaily, M., Hinkle, C., Liu, C., and Bailey, V., 2017,
Shifts in pore connectivity from precipitation versus groundwater rewetting increases soil carbon
loss after drought: Nature Communications, v. 8, p. 1335,
https://doi.org/10.1038/s41467-017-01320-x.
- Smith, P., 2004, Carbon sequestration in croplands: The potential in Europe and the global context:
European Journal of Agronomy, v. 20, p. 229–236, https://doi.org/10.1016/j.eja.2003.08.002.
- Tarnocai, C., Canadell, J.G., Schuur, E.A.G., Kuhry, P., Mazhitova, G., and Zimov, S., 2009, Soil
organic carbon pools in the northern circumpolar permafrost region: Global Biogeochemical Cycles, v.
23, https://doi.org/10.1029/2008GB003327.
- Turetsky, M.R., and 13 others, 2020, Carbon release through abrupt permafrost thaw: Nature
Geoscience, v. 13, p. 138–143, https://doi.org/10.1038/s41561-019-0526-0.
- Veldkamp, E., Schmidt, M., Powers, J.S., and Corre, M.D., 2020, Deforestation and reforestation
impacts on soils in the tropics: Nature Reviews. Earth & Environment, v. 1, p. 590–605,
https://doi.org/10.1038/s43017-020-0091-5.
- Waddington, J., Strack, M., and Greenwood, M., 2010, Toward restoring the net carbon sink function
of degraded peatlands: Short-term response in CO2 exchange to ecosystem-scale
restoration: Journal of Geophysical Research, v. 115, G01008, https://doi.org/10.1029/2009JG001090.
- Walker, T.W.N., Kaiser, C., Strasser, F., Herbold, C.W., Leblans, N.I.W., Woebken, D., Janssens,
I.A., Sigurdsson, B.D., and Richter, A., 2018, Microbial temperature sensitivity and biomass change
explain soil carbon loss with warming: Nature Climate Change, v. 8, p. 885–889,
https://doi.org/10.1038/s41558-018-0259-x.
- Woolf, D., Amonette, J.E., Street-Perrott, F.A., Lehmann, J., and Joseph, S., 2010, Sustainable
biochar to mitigate global climate change: Nature Communications, v. 1, p. 56,
https://doi.org/10.1038/ncomms1053.
- Wu, Y., Ouyang, W., Hao, Z., Lin, C., Liu, H., and Wang, Y., 2018, Assessment of soil erosion
characteristics in response to temperature and precipitation in a freeze-thaw watershed: Geoderma,
v. 328, p. 56–65, https://doi.org/10.1016/j.geoderma.2018.05.007.
- Zhang, H., Wang, E., Zhou, D., Luo, Z., and Zhang, Z., 2016, Rising soil temperature in China and
its potential ecological impact: Scientific Reports, v. 6, 35530, https://doi.org/10.1038/srep35530.