How soil-water flows and how fast it moves solutes are important for plant growth and soil
formation. The relationship describing the partitioning of precipitation, P, into
run-off, Q, and evapotranspiration, ET, is called the water balance.
Q incorporates both surface runoff and subsurface flow components, the latter
chiefly contributing to soil formation. At shorter time intervals, soil-water storage,
S, may change, dS/dt, due to atmosphere-soil water exchange; i.e.,
infiltrating and evaporating water and root uptake. Over sufficiently long time periods,
storage changes are typically neglected (Gentine et al., 2012). Percolation theory from
statistical physics provides a powerful tool for predicting soil formation and plant growth
(Hunt, 2017) by means of modeling soil pore space as networks, rather than continua.
In heterogeneous soils, solute migration typically exhibits non-Gaussian behavior, with
statistical models having long tails in arrival time distributions and velocities decreasing
over time. Theoretical prediction of solute transport via percolation theory that generates
accurate full non-Gaussian arrival time distributions has become possible only recently
(Hunt and Ghanbarian, 2016; Hunt and Sahimi, 2017). A unified framework, based on solute
transport theory, helps predict soil depth as a function of age and infiltration rate (Yu
and Hunt, 2017), soil erosion rates (Yu et al., 2019), chemical weathering (Yu and Hunt,
2018), and plant height and productivity as a function of time and transpiration rates
(Hunt, 2017). Expressing soil depth and plant growth inputs to the crop net primary
productivity, NPP, permits optimization of NPP with respect to the
hydrologic fluxes (Hunt et al., 2020). Some remarkable conclusions also arise from this
theory, such as that globally averaged ET is almost twice Q, and that the
topology of the network guiding soil-water flow provides limitations on solute transport and
chemical weathering. Both plant roots and infiltrating water tend to follow paths of least
resistance, but with differing connectivity properties. Except in arid climates (Yang et
al., 2016), roots tend to be restricted to the thin topsoil, so lateral root distributions
are often considered two-dimensional (2D), and root structures employ hierarchical,
directional organization, speeding transport by avoiding closed loops. In contrast,
infiltrating water (i.e., the subsurface part of Q) tends to follow random paths
(Hunt, 2017) and percolates through the topsoil more deeply, giving rise to
three-dimensional (3D) flow-path structures. The resulting distinct topologies generate
differing nonlinear scaling, which is fractal, between time and distance of solute
transport.
On a bi-logarithmic space-time plot (Hunt, 2017), optimal paths for the different
spatio-temporal scaling laws of root radial extent (RRE) and soil depth,
z, are defined by their radial divergence from the same length and time positions.
RRE relates to NPP, which is a key determinant of crop productivity,
through root fractal dimensionality, df, given by RRE ∝
NPP1/df, with predicted values of df of 1.9 and 2.5
for 2D and 3D patterns, respectively (Hunt and Sahimi, 2017). Basic length/time scales are
given by the fundamental network size (determined from the soil particle size distribution)
and its ratio to mean soil-water flow rate. Yearly average pore-scale flow rates are
determined from climate variables (Yu and Hunt, 2017). Each scaling relationship has a
spread, representing chiefly the range of flow rates as controlled by P and its
partitioning into ET and Q. This conceptual basis makes possible
prediction of the dependence of NPP on the hydrologic fluxes, Q (which
modulates the soil and root depths), and evapotranspiration, given by ET = P −
Q (which modulates RRE).
Consider the steady-state soil depth (Yu and Hunt, 2017),
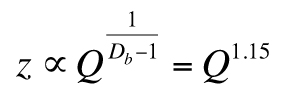 with Db = 1.87, governing solute transport, which is the backbone
fractal dimension of percolation. Optimization of
NPP ∝ RRE ∝ Q1.15(P − Q)df
with respect to Q by setting d(NPP) ⁄
d(Q) = 0 yields ET = P df
⁄(1.15 + df) = 0.623P, within 1–2% of the mean of global
estimates (Hunt et al., 2020).
with Db = 1.87, governing solute transport, which is the backbone
fractal dimension of percolation. Optimization of
NPP ∝ RRE ∝ Q1.15(P − Q)df
with respect to Q by setting d(NPP) ⁄
d(Q) = 0 yields ET = P df
⁄(1.15 + df) = 0.623P, within 1–2% of the mean of global
estimates (Hunt et al., 2020).
The ratio ET/P may be represented using the aridity index, AI, often
defined as PET/P (sometimes as its inverse), with PET being the potential
evapotranspiration (Budyko, 1958). In arid regions, where soil depths are yet increasing,
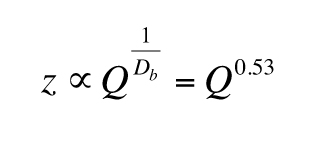 (Yu and Hunt, 2017). For a bare land area, the fraction of the surface that plants occupy
may be only P/PET, which is the inverse of the AI. Both tend to increase
ET as a fraction of P. For high AI, roots are also less confined
near the surface, searching water more deeply, and also increasing ET. Under ideal
conditions of neither energy nor water limitation (AI = 1), Levang-Brilz and
Biondini (2003) determined that for 16 grass and 39 Great Plains forb species the mean
df for all forbs was 2.49, but grasses separated into two distinct
groups with df = 2.65 and 1.67, in accord with percolation predictions
(Hunt and Sahimi, 2017). In the studied biome, grasses constitute more than 90% of the
biomass.
(Yu and Hunt, 2017). For a bare land area, the fraction of the surface that plants occupy
may be only P/PET, which is the inverse of the AI. Both tend to increase
ET as a fraction of P. For high AI, roots are also less confined
near the surface, searching water more deeply, and also increasing ET. Under ideal
conditions of neither energy nor water limitation (AI = 1), Levang-Brilz and
Biondini (2003) determined that for 16 grass and 39 Great Plains forb species the mean
df for all forbs was 2.49, but grasses separated into two distinct
groups with df = 2.65 and 1.67, in accord with percolation predictions
(Hunt and Sahimi, 2017). In the studied biome, grasses constitute more than 90% of the
biomass.
Figure 1 shows our predicted upper bound (dotted line) of ET/P as a function of
AI. At low AI (<1) the known limit ET ≤ PET is
applied. For
large AI, df = 2.5, appropriate for deeper, more isotropic,
root systems. Levang-Brilz and Biondini’s (2003) experimental df values
generate the spread in predicted ET at selected AI values (though
experimental values df > 3 that generate ET > P
are not used). What is new is the representation of predicted variability in ET
based on experimental df value at larger AI, not just
AI = 1.
 Figure 1
Figure 1
Predicted and observed variability of precipitation,
P, and evapotranspiration,
ET,
ET/P, as a function of
PET/P =
AI (aridity index). Data from
Gentine et
al. (2012). Figure is modified from Hunt et al. (2020).
Values of df for grasses generate almost the exact observed variability
in ET/P at AI = 1, but overestimate variability at larger AI. We
attribute the discrepancy at larger AI mostly to the fact that low-end
ET/P values come from grass species with df around 1.9, typical
for nearly 2D structures, being less adapted to arid conditions, and more likely absent at
larger AI. Our theoretical framework, together with experimentally determined
parameters df, generates a good upper bound for ET/P from
theory and its variability as a function of AI.
The most important theoretical limitations of applying percolation theory to water balance
modeling arise from the partitioning of surface run-off and subsurface flow (and
transpiration and interception), because these processes are not obviously regulated by
plants for optimizing NPP. The ability to predict contributions of surface run-off,
plant interception, and subsurface flow would also be important in evaluation of
sequestering carbon and coupling global water and carbon cycles. Incorporating observations
helps estimate these complementary fluxes. We found that variability in the predicted water
balance due to variation in plant root fractal dimensionality outweighs
uncertainties/variation in interception and surface run-off. Coupling our long-term
percolation model with the short-term stochastic infiltration model (e.g., Rodriguez-Iturbe
et al., 1999) might improve predictions of water balance components and optimization of
plant productivity.
References Cited
- Budyko, M.I., 1958, The heat balance of the earth’s surface: Washington, DC, U.S.
Department of Commerce, Weather Bureau.
- Gentine, P., D’Odorico, P., Linter, B.R., Sivandran, G., and Salvucci, G., 2012,
Interdependence of climate, soil, and vegetation as constrained by the Budyko curve:
Geophysical Research Letters, v. 39, L19404, https://doi.org/10.1029/2012GL053492.
- Hunt, A.G., 2017, Spatio-temporal scaling of vegetation growth and soil formation:
Explicit predictions: Vadose Zone Journal, https://doi.org/10.2136/vzj2016.06.0055.
- Hunt, A.G., and Ghanbarian, B., 2016, Percolation theory for solute transport in porous
media: Geochemistry, geomorphology, and carbon cycling: Water Resources Research,
v. 52, p. 7444–7459, https://doi.org/10.1002/2016WR019289.
- Hunt, A.G., and Sahimi, M., 2017, Flow, transport, and reaction in porous media:
Percolation scaling, critical path analysis and effective medium approximation: Reviews
of Geophysics, v. 55, p. 993–1078, https://doi.org/10.1002/2017RG000558.
- Hunt, A.G., Faybishenko, B.A., Ghanbarian, B., Egli, M., and Yu, F., 2020, Predicting
water cycle characteristics from percolation theory and observation: International
Journal of Environmental Research and Public Health, v. 17, no. 3,
p. 734, https://doi.org/10.3390/ijerph17030734.
- Levang-Brilz, N., and Biondini, M.E., 2003, Growth rate, root development and nutrient
uptake of 55 plant species from the Great Plains Grasslands, USA: Plant Ecology,
v. 165, p. 117–144, https://doi.org/10.1023/A:1021469210691.
- Rodriguez-Iturbe, I., Porporato, A., Ridolfi, L., Isham, V., and Cox, D.R., 1999,
Probabilistic modelling of water balance at a point: The role of climate, soil and
vegetation: Proceedings of the Royal Society of London, Series A, v. 455,
p. 3789–3805, https://doi.org/10.1098/rspa.1999.0477.
- Yang, Y., Donohue, R.J., and McVicar, T.R., 2016, Global estimation of effective plant
rooting depth: Implications for hydrological modeling: Water Resources Research,
v. 52, no. 10, https://doi.org/10.1002/2016WR019392.
- Yu, F., and Hunt, A.G., 2017, Predicting soil formation on the basis of
transport-limited chemical weathering: Geomorphology,
https://doi.org/10.1016/j.geomorph.2017.10.027.
- Yu, F., and Hunt, A.G., 2018, Damköhler number input to transport-limited chemical
weathering calculations: ACS Earth & Space Chemistry, v. 1, p. 30–38,
https://doi.org/10.1021/acsearthspacechem.6b00007.
- Yu, F., Hunt, A.G., Egli, M., and Raab, G., 2019, Comparison and contrast in soil depth
evolution for steady-state and stochastic erosion processes: Possible implications for
landslide prediction: Geochemistry, Geophysics, Geosystems, v. 20,
p. 2886–2906, https://doi.org/10.1029/2018GC008125.